|
Check out AI-generated reviews of all Ken Wilber books
TRANSLATE THIS ARTICLE
Integral World: Exploring Theories of Everything
An independent forum for a critical discussion of the integral philosophy of Ken Wilber
 Imre von Soos, architect, civil engineer, research scientist, philosopher and writer is a Hungarian born, Hungarian and Australian national. His anti-communist activities have forced him to escape from Hungary, and he lived and worked since in Australia, France, Germany, Austria, England, Switzerland, Brazil, the Czech Republic and now again in Austria. Read more...
.
The Rational Evolution
of the Species
Imre von Soos
According to the here presented thesis, the Universe being logical, and matter and action by the mind ordered energy, evolution is a self-generated, conscious, rational, mental developmental process of individual elements towards higher stages of differentiation and complexity, higher life-efficiency, higher life-values, more differentiated consciousness and the increasing of the faculties of logic and creativity, resulting in its own continuous geometrical escalation,expressed in continuously restructured abstract and concrete life forms, and is happening in every sphere of the Natural Order. Evolution is in all its manifestations an intellectual affair, and represents for every entity the advancement of its mental power in overcoming its actual psychical and physical limitations. This applies as much to the elementary particles as to the universe as a whole, and signifies that neither is complete in its present state, but is continually evolving. It is anamorphosis: an organism's exercise of its potentialities of evolution to create for and out of itself an improved, new form of life, driven by its intellect-pressure, where survival while a sine qua non is not the prime mover, and where diversity and complexity and not multiplication are the dominant tendencies. The higher the intellect, the more differentiated and expanded is the consciousness, and the higher is the evolutionary state and rate of progress of the individual, who, while cannot control how he is acted on by his environment, is the unique master of his own reactions, and the unique agent for his own change these being the reflections of his individual being , and who readjusts and reorganizes himself constantly according to his external perspicacity and his internal character.
The through self-configuration acquired psychical and physical adaptations are being registered consequently on the genetic material, and transmitted through the reproductive mechanism, meaning that the evolutionary changes happen through and within the maturing and matured entity, that transmits its newly developed qualities to its descendants.
The arguments are founded on the theories expounded in the chapter Natural Order Universal Relativity, according to which Mind is an irreducible and fundamental aspect of Summa Reality; and all "inanimate" and "animate" physical manifestations are the external aspects of the internal integration of differentiated, individuated minds.
The main propositions and empirical rules related to the subject are:
- There exists no concrete boundary between the inorganic and organic realms; and the natural laws applying to one apply also to the other, right through the physico-biological system.
- All life-manifestation has a common origin: the basic physiological processes are characteristic to all living organisms.
- All species and societies are psychologically and physically heterogeneous, their heterogeneity being proportional to the manifestations of individual evolutions within a species, or rather within a society, as species is a biological abstraction. Species evolution represents the mean evolutionary state of the individual evolutions within a species.
- Evolution being individual and gradual, the causes of an intraspecific split, and what appears to be the saltatory evolution into a new species, must lie in the differential rate of individual mental developments. In other words: major evolutionary advances have never represented the collective advance of a whole species, but the natural retardation inherent in the status quo kept by the great majority of a species relative to that fraction which continued on naturally advancing in their particular evolution, representing first a sub-species, then turning finally, through an intraspecific split-off, into a new, more complex and more highly organized species.
- The relative regression of one part of the species, and the liberated expansion of its evolution-vital fraction into a new species, gives the impression of an evolutionary jump.
- The evolutionary split is the consequence of a continuous genesis rhythmically bursting into wider expressions of speciations.
- As the evolution of a physical organ must be contemplated in the light of how that particular evolution has promoted the evolution of the organism it forms part of, so the evolution of every fraction must be regarded as the differential evolution of the whole, forming an interrelated open system, where each is the subject, instrument and originator of Summa Creation, that happens every time the consciousness of the fraction transcends itself.
A further proposition of fundamental importance is, that neither the single cells as unicellular elements (protozoa), or as constituting elements of multi-cellular organisms nor the multi-cellular organisms (metazoa) as such have reached their peak or their dead end in evolution; and that their evolutionary criterion lies not in the comparison to the highest evolutionary forms, but in the efficiency with which they fill their respective ecology-niches in the Planetary Organism as subjects, instruments and originators of Evolution; a criterion that defines their universal life value.
Figure 14-1 shows the biological classifications and the simplified direct line of evolution culminating in Homo sapiens, with two question-marks into the future, that the present thesis also attempts to answer according to the apparent evolutionary rules. There are several facts empirically obtained to be added to the above diagram, and others that can be read from it.
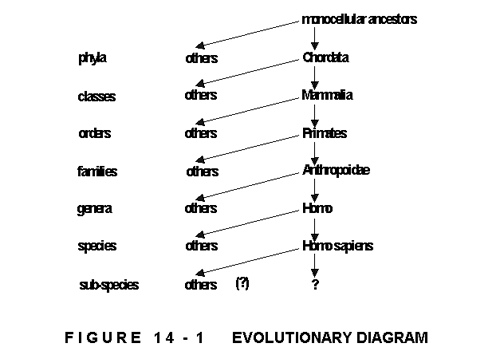
In the history of evolution, once the subdivisions have established themselves within a certain division (e.g. classes within a phylum, orders within a class etc.), no more subdivisions have developed within that division. The very same number of phyla existing today, existed already after the sudden diversification, called the Cambrian Explosion about 600 million years ago at the beginning of the Palaeozoic era. Within the phylum Chordata all classes were established 250 million years back. All the Mammalian orders between them the Primates have existed for the last 100 million years. The last Primate family, the Anthropoidae, were established 25 million years ago. The timing for further subdivisions is uncertain, while species and sub-species are still in differentiation, and so might be, or rather is following the universal rules Homo sapiens.
Another remarkable fact is, that major advances were made linearly, progressing through the subdivisions without crossing, meaning that the most advanced species came from the most advanced genus of the most advanced family of the most advanced order of the most advanced class of the most advanced phylum. The same applies right through the system. This points to the fundamental truth that differentiation, that is, evolution of new sub-types does not come about by random chance, but by intelligent self-design, where the more evolved type has the better means for further development: its brains. The process, being self-reinforcing, is exponential. It also means that intraspecific differentiation starts with the differentiation of individual mental faculties within the species.
Figure 14-1 shows the differentiations along the most advanced evolutionary process on this planet. This, however, most emphatically does not mean that the 'others' have reached a dead end, stagnation, or extinction. To believe that only Homo sapiens evolves, is but an anthropocentric, self-induced and self-deceiving preconception. Neither, just as emphatically, does it mean that these 'others' become naturally subordinated to the 'leader' who becomes a ruler and serve as 'free game' for his pleasure and abuse. These 'others' also evolve, but at variously slower rates, and fill their respective niches as indispensable symbionts in a balanced ecology an evolving Planetary Organism , an act the self-appointed 'all mighty leader' apparently fails to do.
The process of intraspecific differentiation can be illustrated with the results of some experiments in heredity with laboratory animals, where random groups were tested for intelligence, divided into a 'dull' and 'bright' group, which were then interbred separately, and compared at every generation. Figure 14-2 shows the Gaussic distribution curves indicating the results, projected unto a three-dimensional graph, the x, y and z axes representing intelligence quotient, time (generations) and frequency respectively.
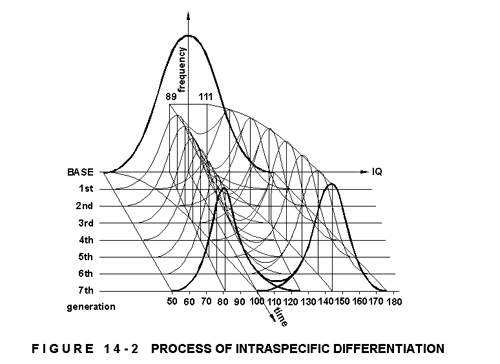
After the seventh generation there were no noteworthy alterations in either group. The IQ calibration is arbitrarily chosen for comparative analysis, the mean of the original group taken as 100. The distribution or standard deviation curve is constructed according to the general statistical rule of distribution, where the central third of the range is occupied by 68,26% of the group, the bordering 1/6th on each side by 13,58% each, and the extreme 1/6th by 2,14% each, leaving 0,14% below and above the range. The point of inflexion that marks the greatest frequency variation is at the quarter points. The central parts of the range (from -1,5SD to +1,5SD) contains 86,1% of the group, the lower and upper extremes 6,95% each.
Intelligence, while no doubt transmitted genetically (organic transmission), is increased only through the active use of this potential. It cannot be learned. The environmental influence comes only in the form of a challenge, to which the self-generated and self-activated reaction means the exercise and consequent increase of the intellectual faculties. This is an individual achievement producing the corresponding inner satisfaction, and stimulating the further use of these individual intellectual faculties, and the acting according to them. Follows that intelligence and achievement (achievement, and not social or financial success!) are inseparable. In opposition to this are the socially and educationally conditioned automatic reactions, the resorting to which are intellect-dulling, and produce the feeling of lack of achievement, frustration and even lethargy.
The drive to exercise the intellect that has its motive and end in itself, and which I am calling intellect pressure has various names in psychology, like 'stimulus need', 'variability need', 'psycho-logical need', and is found even in the lowest organisms. The drive, the exercise and the intellect are interrelated, which is why the bright gets brighter and the dull duller. There is still another factor that is a function of the intellect: the stimulus complexity. To every intellect level belongs a corresponding spectrum of complexities, below which the mind is disinterested, above which confused. Towards the upper limits of the spectrum are the spacer stimuli, providing the challenge, that, once mastered, cause the spectrum move towards higher complexities. This complexity spectrum defines the intellect-level of the individual, while the spacer-stimuli, that represent his intellect-pressure, are acting on his most vital functions, those which produce his syntropy.
Founded on these same lines of thoughts, organic evolution characterized by growing complexity and change works through each differentiated fraction; through the particular effort of each individual being, to react positively to the challenge of the change and growing complexity created by through all its fractions evolving environment external induction , but mostly through its own "intellect-pressure" acting as its evolutionary drive internal induction , thus becoming itself more complex and with it contributing to the general evolutionary flow. It is self-generated and self-activated. So are regression and degeneration. Evolutionary history is the history of the evolution of individuals as such, and of the prevalence of the perpetuation through descendants of evolved individuals of a species, forming first a new subspecies through intra-specific differentiation and finally turning into a new species through evolutionary split of a higher evolutionary state: a super-race in relation to the one from which it evolved. In this intellectual and spiritual endeavour the individual is not selfishly self-managing against the others, but is self-transcending for its own and consequently for universal evolution. The misrepresented portraying of this absolutely natural life-manifestation of "everyone by himself", prevailing on this planet for three and a half billion years, as "the egocentric ideal of attaining egocentrically the extremity of 'everyone for himself'", might caress some primitive archicortical crannies, but cannot reach the neocortex, and the mind especially not.
The whole evolutionary drive is directed towards a higher aim, higher form of self-expression, towards the realization of the self on a higher plane, which transcends the actual level of its being. It is the expansion of spirit towards higher realizations, and consciousness towards greater realities: this is self-transcendence.
Within the evolutionary drive of an individual belongs his self-assertive tendency, which comes to manifestation on the individual level, and against other elements of the same kind and function, through the intra-specific competition for the most suitable type to fulfil that particular function. It is in accordance with the law of the survival of the fittest or rather the prevalence of the most able , and is basically in its natural form for something in a higher order, and not against someone in the same, where the success of the superior individual is at the same time through its effects the success of the entire system.
When the base-group (mean IQ100) was separated into two, the 'dull' and 'bright' groups had mean IQ-s of 89 and 111 respectively. By the seventh generation the 'dull' group regressed to 81, while the 'bright' one progressed to 143. The relationship being logarithmic, the bright group's final mean was four times superior. Only the top 6,5% of the 'dull' group was brighter than the dullest of the 'bright' group, and 6,5% of the 'bright' group duller than the brightest of the 'dull' group. So, by the seventh generation the intellectual differentiation was from any practical point of view complete. It is noteworthy, however, that 15% of the first generation of the 'bright group was below the original mean (IQ100, which was the lowest selected parent), diminishing to zero by the seventh, while 35% of the 'dull' group was above this figure, as against the final 6,5%. As all this cannot be due to genetic transmission only, it points to the spacer stimuli being situated at both ends of the complexity spectrum, and influenced by the general intelligence level and tendency of the group, thus clustering and separating peak values, and accounting for the arrival of the fittest on one end, and the departure of the unfit on the other. This phenomenon is well noticeable in the actual human societies.
The selections and the conditions that brought about the above results were laboratory selections and conditions, and their results could be claimed to be consequently unrealistic. However, similar kind of selections, based on abilities corresponding to particular species environments, are going on constantly right through the living Nature, brought out by population pressures and manifested through the mechanisms of decisions by proving superior relevant abilities, pecking rights, the unfit progeny's culling by the parents, falling to predators, or general incompetence to survive, where the parents' aim is not towards creating a 'better future' for a whatever progeny, but a better progeny to be able to face a whatever future. It has to be stressed, that evolution happens basically on individual level, and is enhanced, beside the particular intellect-pressure, by the evolution of the other individuals forming the group, by the surrounding predator and prey evolution, and by the manifold environmental adversity. The mechanism for the consolidation of the newly gained positive characteristics, promoting species improvement, complexity and diversification, is the reproductive mechanism combined with the mating rights of the superior males, all representing in practice the "reproductive isolation" of a superior subspecies.
Evidently, the higher becomes the mean intelligence within a species, the broader becomes the intraspecific intellectual spectrum, and the greater become the differences between the particular psychical environments that the various individuals of the same species, and even of the same species populations live in, even if the physical one seems to be the same for everyone, and just as evidently the greater becomes the psycho-physical and energetic heterogeneity of that species, resulting in a corresponding intraspecific diversity in the rate, trend, progress, complexity and the amount and nature of branching. The differentiation, especially in its developmental stage, is psychological, marked by little or no morphological deviation, expressing itself also morphologically only after the consolidation into the new species, as the physical adjustment to the psychological changes. This latter might well be the explanation for the observed "punctuated" morphological changes in the fossil records following a gradual psychological evolution, manifested in an "evolutionary jump". And the second reason for the quasi non-existence of the missing links might be, that the new species represents always only a fraction of the one it evolved from, and expands in numbers only after the concretisation of the "evolutionary split".
That a cataclysmic occurrence can split a species geographically is imaginable; that this separation happens into dull and bright groups, is improbable. The thus separated groups have to adapt at least their feeding habits, and to some degree to their new and diverse environments, resulting in some morphological adjustments, thus differentiating from each other and from their original state. This process could produce, depending on the severity of the new environmental challenge, and the ability of the individuals of the groups to react to it, two new sub-types, one or two branching-offs, but it couldn't produce an "evolutionary jump" resulting in a higher evolved new species.
Environmental changes do not refer only to climatic conditions, but very much also to dynamic readjustments within the ecological communities, where particular evolutions force out constant changes in the feeding interrelations and conditions of survival. Both Darwin and Wallace have pointed out "the inevitability of evolutionary change in a world in which each species tends to produce more individuals than can find a place, provided that there is sufficient variability, at least part of which is transmissible to the next generation." While some general species variations occur as adjustments and adaptations to environmental changes be those climatic, biological or ecological their motivation to self-improvement is concentrated on survival. On the other hand, behind the particular evolutions is the already discussed intellect-pressure, the evolutionary drive of individual members of the species, that is self-generated without any external induction. Intellect-pressure works as a drive for self-enhancement, rendering it the primary cause of evolution. This is also reinforced by the fact that the higher is the evolutionary level of a species, the faster is its evolution, and the shorter are the intervals between the evolutionary jumps producing a more evolved species; a fact that also counters the theory of random mutation as the cause of evolution, which, being random, should work with equal timing at all evolutionary levels. Both variations are working on the trial and error principle, like all creative and productive endeavours do; the trials are, however, not chance or random occurrences of ad hoc genetical mutations unfolding within a maze of labelled but undefined selective agents, but self-generated, conscious and intelligently elaborated initiatives or reactions, dynamically readjusted during the process: chess game with fixed rules and flexible strategies, as against the passively hoping for some dices to be falling in one's favour. This later happens to be the ossified credo of materialist biology, in keeping with the ad hoc universe of materialist cosmology.
My theory that evolution and thus intraspecific differentiation happens on intellectual level, has its foundation in the aforesaid, and shall be elaborated further as this argument progresses.
As most of the life-manifestations and attributes, like self-preservation, feeding, reproduction, acquisitiveness and even leadership and constructiveness are ascribed by materialistic science to some extraneously implanted pre-programming, called 'instinct', let us indulge first in a closer scrutiny of that concept.
INSTINCT is one of the labels with as many meanings as there are users, to interpret animal behaviour and to excuse the antics of Homo sapiens. Until recently even prominent scientists believed (some maybe still do) that animals this time excluding man are biological machines, without consciousness, intellect, or any thinking and reasoning capacity, functioning only mechanically according to their 'instincts'. Neither the meaning of this word nor the origin of the concept was important as long as it has served for a label that requires no further explanation. It became the cause of innumerable effects, without itself having its cause determined. The machine-theory has ceased the fogging not.
Dictionaries define the word 'instinct' as "innate propensity to certain seemingly rational acts performed without conscious design"; "innate impulse"; "unconscious skill"; "unlearned complex adaptive response"; "the original sense"; "congenital impulse plus specific emotional excitement". Unless accepting the edict that order derives from chaos thorough random action, "without conscious design", none of the above definitions make any sense: they are labels without substance. One should wonder at the coincidence that 'unconscious nest-building skill' has happened to birds and not to elephants; or maybe it has happened to both, the latter 'instinctively' rejecting it for not having 'survival value'. Or maybe those who are still labouring over the chronological order of egg and hen, are just simply not asking the right questions.
Rejecting acausality in the case of a fundamental cause of many life-manifestations, I will follow the appearance and development of instinct as an integral part of the appearance and development of biological life-manifestation itself.
While it would be possible to go further back, a more understandable point of beginning is at the unicellular organisms, the protozoa. All cells have the potentiality for movement at will. Not all move. Those that do, had to learn it step by step, by trial and error, until co-ordinated movement was achieved, and, through repetition, changed from consciously directed action to motor action, to be called up as one unit of unconscious process routine by one stimulus, in this case the will. The will to move was conscious, the twitchings co-ordinated into movement became instinctive. Forward, backward, left, right up and down, each had its own program pack (software), to be triggered into action by the correct stimulus (code).
Sensing food or danger, the cells willed their movement in the appropriate direction, learning to respond correctly. These actions became routines through repetition, automatic actions, triggered off directly by a sense-stimulus. 'Approach' and 'withdrawal' had the capacity to follow a preset course and strategy, and activate the correct movement, without its details having to enter into consciousness.
The cell had now two new programs for reaction, both using the already existing movement sub-routines, which remained nevertheless still accessible to the conscious will. Delegating routine work to a sub-system to what is generally called an "unconscious motor action" the cell became free to evolve ways of further development. The programs recorded on the chromosomes joined the sets of informations which guide the physiological activities of the organism. When dividing, the acquired behaviour patterns were transferred, became innate in both new parts.
Based on the foregoing, instinct can be defined as fixed pattern of mental or physical behaviour, innate or acquired, in living organisms, triggered off by specific, simple stimuli.
As is explicit in the above study, instinct, or rather the acquisition of them, even in the simplest manifestations of life, are preceded by a conscious cognitive process (programming) expressed by the thinking of aspects, qualities and relations, exercising comparison, generalization, abstraction and reasoning.
Instincts are also characterized by their fixation, which depends on two factors:
Firstly, how far are they removed in the hierarchic order of mentation, how deep they are in the program memory, to what extent did they remain directly accessible to conscious will.
Secondly, how powerful was the original motivation, and what is the frequency of its use.
To be able to erase, modify or reorganize a fixed behaviour pattern, its origin has to be found by regression, recognized in its whole run, and attended to step by step with the same force and motivation, as the original input had. There is an inverse relationship between the intensity of the drive to instinctive behaviour, and the level of creative intellect of the individual. The same relationship exists between the innate/acquired factor of instinct and the same level.
I have to emphasize that behaviour, rational or instinctive, irrational or random, can happen on the individual level only. While a collective average of racial characteristics can be spoken of, collective intellect or collective instinct never. Herd-conduct, tribalism and nationalism are the mass-expressions of elementary units of conditioned behaviour, and are not to be confused with the self-orchestrated rational behaviour of heterogeneous elements in natural social conduct directed towards the symbiotic development of an integrated organism: an individual of higher complexity and identity, the evolution of which is integral part of the universal evolution. Such behaviour aggregated macromolecules into organelles, organized them functionally into cells, differentiated the cells into specialized elements, and produced, through movement, cohesion and co-ordination, the plant and animal organisms, and joined them into major ecological communities.
The only true multi-organismic societies are the insect societies, but they can hardly serve as examples, for they are one-family societies, meaning that all their specialized members, as also the perpetuation of the species, arrive out of the one single female, the queen, who controls through this fact the equilibrium, advance, social processes and population growth of her group, eliminating ab ovo any possibility of bifurcation, or even the smallest evolutionary split. It is even questionable, if these should be considered as societies, or as organisms. While all termites and ants are social, only 8% of the wasps and 5% of the bees form societies, the rest are solitary, but definitely not being inferior for that, possessing even superior evolutionary possibilities.
Insects and lower vertebrates, with some exception, do not know their parents, neither do they grow up in a community of various age-groups, consequently are born with their inherited ready mechanism serving for every manifestation of their life. Their mental activities consist of making decisions according to their fears, needs and desires, and activate corresponding programs with fixed rules and flexible strategies. These animals are born free: independent; they do not depend on anybody, only their own brain and strength for survival. Innate instincts being rigid, the evolution of the species through its individuals is slow, if not stagnant, although the physiological intelligence of some insects, making adaptations to before never encountered variations in the environment, which are then genetically transmitted to their progenies, is more than amazing. However, these changes are adaptive and not evolutive.
The variety in the animal kingdom is enormous. Most birds are as helpless when born as primates are; totally dependent. Flying and obtaining food is taught to them by their parents. If hatched in captivity they might acquire both as autodidacts, but their capacity to survive will be much inferior. Paring rituals and nest-building techniques are instinctive in the sense that they are oriented by innate knowledge.
Anybody who is watching with the eyes of a participant, the first wing-probes of young birds, and not only sees but feels emanating the fears and desires to fly, the loving, strategic encouragement and impatient disapproval, the anxieties and the pride, could never consider this scene as 'biological mechanisms' being watched by a superior creature. This human prejudice, born of ignorance and supported by arrogance, is also a conditioned response, where the above mentioned instinct/intellect relationship applies.
The hoofed mammals, when born, can move freely, but depend for food and protection on their mother. The co-ordinated body-movement is certainly inborn; if their general pattern of behaviour, typical to the species, is governed by instinctive-innate or instinctive-learned forces, or by the mixture of the two which is the most likely will not be known with absolute certainty until we can tune in on their communication system. Until then we'll have to be content with the already mentioned waste material passed out of the alimentary canal of Waddington's cow at best , or of that of her male partner at worst.
One of the animal behaviours, or rather behaviour complexes I have always most admired, is the aptitude test and its follow up in connection with mating and procreation. This probe of courage, ability, strength and endurance not an angry fight! decides which one male will be the leader responsible for the well-being of a herd of twenty to forty females, who will also be the one and only inseminator for that year, and for each year he can maintain his position by proving his aptitude. The implication of this is stupendous, and not only in the assurance of healthy progenies. To accept a 'man to man' test, adhere to its rules, and accept its decisions, indicate a moral standard that is reminiscent of the knights of the round table, or of the golden age Aurea which sponte sua sine lege fidem rectumque colebat. There is no ganging up by the losers against the winner, no cloak and dagger, no calling for some cock-eyed 'justice' to share and share alike. The moral strength of each looser is greater than its sex-drive, its self-assertive tendencies, its want to dominate.
Should your answer be that these unfortunate creatures have an "irresistible inner force compelling the performance of this act, without, or even against the will of the individual performing it", consider that at one stage in the evolution of this animal, every instinct was a consciously unfolding act, which became mechanical only by choice and repetition. No behaviour pattern could have evolved if it was not self-generated in the interest of the individual who lives in physical and mental communion with the whole.
The carnivores of the dry land are born blind and are fully dependent. They live in families or in family groups, where the young are individually or collectively taught both their behaviour and their hunting habits.
Two thoughts appear to me interesting in their relation:
Many kinds of the carnivores in the wild do not procreate if a bad year (e.g. drought) is in coming. This foresight cannot be instinctive pre-programmed , only intuitive psychically perceived , and throws light onto another intelligent, healthy animal reaction: procreate only if the progeny has good chances for a normal, healthy life. In other words: they create neither runts nor misery!
Connected to this fact is the second observation: predators keep their numbers constant, adjusted to the number of the herbivores whose old, weak and sick they feed on, thus keeping also their numbers unaltered, and the herd healthy and strong. Thinking as humans do, they could multiply themselves as there is enough food for two or three generations to come, until the herbivores disappear and 'après moi le deluge'. Or they could at least continue reproducing in the first case, as they are only indirectly influenced by the drought. They do neither.
The reason cannot be the favourite reply: instinct, which is destined to fit every occasion but explains none. The race could not have pre-programmed itself for an 'automatic reaction' to be triggered off by a psychic perception in the first case, or at every mating season by the result of the counting of the respective numbers, in the second. For this not only the foresight of the coming season, but of the repercussions of the present action on the life of two generations ahead, would have been necessary, a feat to which humanity most certainly is incapable. It is more likely that they answer positively to the co-ordinating plan of the ecological community which they form part of and are in conscious harmony with.
Highest on the list are the primates, whose progenies are completely dependent on their parents not only for a short period after their birth, but for the first 15 to 20% of their life-spans. From the co-ordinated action of their muscles to reach for food, to social behaviour and skill to survive, every step, every action has to be acquired. This opens tremendous possibilities for evolution, and tremendous dangers for abuse, depending on the quality of the immediate environment. The dependent infant being also highly impressionable becomes the product of the group-life, society; his personality and its formation, and his social attitudes, are formed by those of the group, communications, interpersonal relations, crowd and mass behaviour and social movements.
With animals in their natural habitat everything is directed towards survival and evolutionary progress. All instincts are natural and are created in this spirit. Aberrations simply do not survive.
Man has created his own environment, removed it further and further from what would be a natural habitat, and has made mental adjustments to his unnatural surroundings. Instead of producing natural instincts by creative, intelligent reasoning process, acquires habits, prejudices, preconceptions, complexes, inhibitions born of ignorance, conditioning, indoctrination and general brain-washing, these feeding on their own products by making continuous loops.
"Where instinct predominates says C. G. Jung psychoid processes set in which pertain to the sphere of the unconscious as elements incapable of consciousness." I am adapting this statement to the terms of this text: "When lower, fossilized systems dominate the higher ones, psychoid processes set in, the control of which pertains to the sphere of the lower systems incapable to be reached by the individual's consciousness and will."
Now back to evolution.
Following the above thoughts, two kinds of intraspecific differentiations can be established:
- by adaptive pressure induced adjustments to environmental changes expressed in minor alterations of feeding, self-protecting and other general behaviour and habits, and corresponding physiological and morphological adaptations;
- by intellect pressure induced manifest evolutionary progress, accompanied by psychological, physiological and morphological modifications.
There is a mental process in both cases that precedes and accompanies the change, that consists of analysing and considering the actual situation, defining new aims, studying possible alternatives of the courses of action and their probable consequences, making a logical choice, and willing it into execution. These are functions of the individual intellect.
In the first case the process is brought about by external circumstances, where, the externally set challenge once met and the adjustments accomplished, the process stops, existence continues in the newly established pathway, the corresponding physiological and morphological adjustments are marked on the chromosomes, the new qualities transmitted through the reproductive mechanism, and consolidated through continuous routine for survival. It is very questionable, however, if these changes would represent an evolutionary progress in relation to the previous state and the rest of the species, or only an intraspecific, "low energy" variation. The adjustment could even be regressive, if, for example, it has involved a greater degree of specialization, or the morphological changes restrict the spectrum of further development possibilities. Neither the quantitative division into geographical specializations, nor the intergradation of different species is an evolutionary step, because specialization which is regressive means not diversification which is progressive and intergradation even if it is successful produces mostly reproductively and intellectually sterile hybrids, but never superior species or life-forms. And neither is the growing from a small and stupid organism into a big and stupid organism a progress, nor is it the explosion of a small and stupid group into a big and stupid one. Neither sizes nor numbers are magical: only quality is.
The overwhelming majority of the morphological differences by which species of the same lineage can be distinguished, can be attributed to adaptive pressure.
In the second case the originative agent is mental: intellect pressure, a self-generated, creative intellect-dependent cause, defining its own objective, its own spacer stimulus. Once the challenge is mastered, the process does not stop: the intellect pressure continuously sets new spacer stimuli with increasing complexity, rising with it in turn the intellect-level also continuously, rendering the escalation exponential. Being self-reinforcing, it is a "high energy" process. However, its transmission has congenital value only if it is further applied and generated, and thus, the efforts not being for adaptive self-preservation but for self-expression and widening horizons, its spreading is limited. The corresponding psychological, physiological and morphological modifications are manifested in the expansion of the nervous system including brain-capacity, less muscular strength but higher muscular skill, lighter general structure with more flexibility, and the diversification of psychical and physical behaviour. As evolution is marked on its whole spectrum with growing cephalization and diversification, the process is decisively evolutionary.
While the first case may account for slow and reasonably homogeneous species improvements with gradual transformations along direct courses, manifesting minor sub-specific adaptive variations, it does not account for the richly branched evolutionary tree with ever increasing complexity of organization. Only the second case can be called for the explanation of the great evolutionary differences and what appear to be "evolutionary jumps", but even that only with the support of another phenomenon, called the physiological intelligence of organ systems, organs, tissues and cells, supporting in harmonious co-operation the activities of the whole organismic apparatus.
For the deeper understanding of this statement, another study is in order.
The cell the basic building stone of all organic life-manifestations had terminated its major development in two slightly different forms: plant-cells (protophyta) and animal-cells (protozoa), about one billion years ago. While its general conception was complete, it still could improve the technologies of its metabolic processes according to its particular functions, and raise its general economy and efficiency. It has not reached the peak or the dead end of its evolution; on the contrary: due to the cell's creative intellect exercised on its own level in response to organismic demands, are the developmental potentials of multi-cellular organisms essentially unlimited. There is a very high degree of order and logic in cellular structures and functions, that reflect on and apply to all biological systems. In fact, function and structure are interdependent and inseparable, function defining structure which is its organ of expression (a statement that has brought me innumerable arguments in architecture and engineering). Evolutionary change itself means a functional modification that requires consequent structural adaptations. All technological changes, from the development of sexual reproduction, the four-chamber heart and the three-cortical brain, to the development of the microscope, telescope, the automobile the aeroplane and the computer, are structural adaptations and extensions to serve functional requirements, these in turn being dependent on mental development.
That the emergence of novel features of increased complexity evolution prevails already right through the inorganic realm, has been discussed earlier in this book. How many evolutionary changes it took for the cell to form out of its basic inorganic elements, I do not know; the important fact to know in this context is, that there exists no concrete boundary between the so called inorganic and organic realms; that the natural laws applying to one apply also to the other; and that self-formed and very complex physicochemical systems (carbohydrates, lipids, proteins and also RNA and DNA), constructed mostly of amino-acids, with self-duplicating property and capable of breaking down others and reorganizing their parts according to their own patterns (representing the beginning of predation) have aggregated into specific and dynamic clusters, symbiotic associations, each capable of carrying out one of the steps in systems of necessary components from available substances, called organelles, which structured themselves into cells. Neither the organelles are yet really understood, nor the constituents, mitochondria; this latter, while being a vital symbiont supplying enzymes essential in catabolism, lives its own life in the cytoplasm, and has its own genetic material, independent of that of the cell. Cells, as individual organisms, are ubiquitous and form the phyla of protozoa and protophyta.
A further step in the development is to stay attached division after division, the thus increased mass and co-ordinated movement being advantageous to each autonomous individual. Specialization on cellular level turns this non-specialized aggregation into a cell-colony, where differentiated groups of cells tissues perform specific functions, and thus create a co-operative society. It has to be remembered that all members are the outcome of the cleavage of one zygote, and that any of them can detach itself and start a new society, that will then grow into an identically differentiated and constituted society like the original one. These specialized cell-colonies are called parazoa, and the sponges are well-known examples of them.
Further differentiation and integration results in a new biological concept: the multi-cellular organisms the metazoa where the cell can no longer separate itself and live outside of it, but neither is the organism a whole without each kind of constituting cell, for there is some special contribution, which this and no other can make most effectively to the life of a rational organism. The relationship and differentiated mind-identity on every stratum was exhaustively discussed when talking about holons.
The first step in the metazoan development was the formation of a food-cavity in the shape of a cup, consisting of an inner layer of cells the endoderm , a second germinal layer developed by and out of the original ectoderm, to be concerned with the alimentation of the system, while the ectoderm can, as before, be concerned with the environment, and the development of the system. The phylum Coelenterata e.g. jellyfish has this type of construction.
In the next move the ectoderm established a third germinal layer between itself and the endoderm: the mesoderm, and with it some primitive structural organs, where organ-formation involves differential cell-migration, differential mitotic rates, pattern formation and sub-cellular changes. These are the Platyhelminthes, the flat-worms.
A further step improved the organs and developed a digestive tube and a vascular system with a cardiac tube, both very primitive, creating the phylum Nematoda, the round-worms.
Each of these steps has opened up possibilities to ever greater diversifications, characterized by increasing grade of complexity, organization and efficiency.
The line of evolution towards Homo sapiens took the steps through the phylum Echinoderma (e.g. sea-cucumber), into Chordata, the early Vertebrates, Lungfish, Amphibians, Mammals, Primates, Anthropoids, Homo, and actually, but not finally, Homo sapiens.
The animal zygote is a monocell structured as above, differing from the protozoa and from each other only by the genetic material (Chromosomes) contained in their nuclei. In the embryonic development it goes first through seven cleavage divisions (128 cells) forming a cell colony called blastula, then comes a multi-cellular organism by forming the ectoderm and endoderm, creating a primitive food-cavity (blastulation), followed by the third germinal layer (mesoderm), and the subsequent organ formation through histological changes. The endoderm turns into the elaborate digestive system, the visceral organs; the ectoderm transforms into skin, sense organs, and the nervous system; and the mesoderm into the skeleton, muscles and vascular system. The unfolding follows the evolutionary development of each singular organ. The heart, for instance, is first a cardiac tube, which changes, following all intermediate steps, into the four-chambered complex. The human embryo looks at 18 days like the amphibian one, and at 27 exhibits bronchial bars reminiscent of the lung-fish ancestor. The transformation of these gill-arches involves a most remarkable rearrangement of skeletal elements, muscles, blood-vessels and nerves, characteristic to amphibian metamorphosis. The muscles are first laid down in a regular segmental arrangement as are in the adult forms of the primitive vertebrates, and subsequently rearranged to form the characteristic arrangement of a human adult. A tail is developed, then withdrawn; an example of the transitory traces of abandoned structural elements.
The development of the embryonic body is remarkably alike in all classes of vertebrates, resulting in a common structural pattern. The internal anatomy and embryonic development of the anthropoid apes are in very close agreement to those of Homo sapiens, as are their menstrual cycle, period of gestation, placental formation, blood, chemical similarity, the only differences being morphological and in the size of the brain, representing a genetic deviation of one per cent.
All life has a common origin; the basic physiological processes are characteristic to all living organisms; all flora and fauna, including man, are related by descent; the differences arising through splitting, branching off, i.e. part of the species making an "evolutionary jump", while the rest maintain their status quo. The branching off has created the evolutionary stages, and its particular history is written in the ontogeny of every organism. This is expressed in Ernst Haeckel's biogenetic law, that states that "Ontogeny, the development of an individual animal is a shortened recapitulation of phylogeny, the evolutionary history of the species to which it belongs."
That all life has a common origin, that the basic physiological processes are characteristic to all living organisms, and that all planetary biological life, including man, are related by descent, are not theories but rationally irrefutable scientific facts. One of its watertight proofs is also expressed in the above quoted Haeckel's biogenetic law. On the other hand, the idea, that the qualitative intraspecific segregations have come about from one part of a species making an "evolutionary jump", while the rest maintaining their status quo, is one of the theories, attempting to explain the apparently abrupt origin of new species during the evolutionary history, and the missing of the transitional links the "missing links" palaeontology fails to come up with all pointing to an abrupt jump-like origin of a new species. I have to point out here another observational fact, that the lower the evolutionary state of a species, the slower is its evolution; and that some species have arrived already over six hundred million years ago to such an extreme stability and quasi-finality in their habits, structures and specializations, that has left no opening for them to any further variation. These individuals had deflected from the pioneers' gradient due to their inertia at an early stage, and the intellectual spectrum and general specialization of their species became so narrow, and the scope of individual improvement or chance so small, that only the slightest, gradual transformations and diversifications were possible. Consequently no particular point exists where one kind of developmental stage ends and the other begins.
Biological science is labouring for centuries to answer two main questions: (1) why did one part of a species make an "evolutionary jump" leaving the rest behind; or much more correctly: why did sub-species within the various species develop higher evolutionary values, which have led finally to "evolutionary splits"? and (2) how, by what kind of biological mechanism, did the split materialize?
There are diverse general lines of thoughts and theories on creation, evolution and evolutionary differentiation, in various degrees of ossification. Negative approaches, dogmatism, materialism and probabilistic mechanicism, all seen through reductionist visors, occupy practically the whole spectrum of thoughts, of which here are some for tasting.
According to Sigmund Freud, evolution is the derivative of continuously frustrated regression towards protoplasmic existence; an idea to which I have no desire to produce any comment. Another negative approach holds, that new breakthroughs are desperate escapes forced out by extinction-menacing dead ends.
Most scientists, forced by their social environment and shackled by their particular specializations, are producing theories within their set limits. Sadly, the social environment can be so powerful, that the so called "Creationists", utterly disregarding the empirical facts presented by astrophysics, geophysics, palaeontology, biology and all the other related sciences, can decree (unpunished!) Moses' Genesis to be the one and only authority on the subject, and the uniquely true presentation of Reality supplying a clear answer to both the "why" and the "how". As a matter of fact, less than half of the Americans would accept that humans descended from other species, and the rest is so adamant as to insist to have exclusively this brainy variant being taught in public schools, as it also is in many colleges of the United States, in spite of contrary Supreme Court rulings. Not being based on any scientific observation, but even contradicted by all, it is a hermetically closed dogmatic belief system, not open to any discussion. I have no knowledge how they reconcile these teachings with the rest of the sciences in these colleges, and how their students fare in their later life-work.
The actually strongest line is represented by materialist science, reigning the greater part of the biological community calling themselves neo-Darwinists, who explain Evolution with probabilistic ad hoc occurrences one could characterize as "micro-bangs". How Darwin's name got into this credo, is beyond me, for Darwin never was a materialist, and, the genius that he was, has presented his theories as open propositions, never allowing them to congeal into dogmas. His supposed followers who are not the first ones to use a great being's name to reinforce with it their rickety credo explain both the "why" and the "how" with accidental chromosomal mutations. Sticking to materialism and acausality as their fundamental premise, they are defining the mechanism of an ad hoc phenomenon, instead of the driving spirit of a rational evolution. Their accidental incident happens to derive from chance-mutations in the genetic material of the nucleated cells, coupled with subsequent "selection" according to some norms, acting jointly as causeless originative agents, with evolution being their lucky product. While their starting point is the nucleated cell, treated as a predetermined mechanism, their theories do not cover the why-s and how-s of its most remarkable development, neither the emergence of any of its manifold constituents in an environment consisting originally exclusively of methane, ammonia and water. And even further: neither does their theory consider, that the emergence of novel features evolution prevails already right through the inorganic realm, without any concrete boundary existing between it and the organic one; and that the natural laws applying to one apply also to the other. They voluntarily ignore, that random chromosomal mutations would lead primarily to the appearance of life-incompetent freaks on the whole spectrum of species evolution not only on that of Homo sapiens and not to improved genotypes representing evolutionary progress and speciation; that even the positive mutations would produce a great spectrum of incompatible varieties, resulting in degenerative or lethal congenital characteristics, and not in life-competent groups of individuals spearheading the evolution of species; and further, that under the rule of randomness, advances would also be random, times one species getting ahead, times another, major advances criss-crossing each other through the subdivisions, and not made linearly without crossing, which is decidedly the case. Their theory also ignores, that this kind of "selection" would tend at best towards the mean, towards the best suited for the particular environment, representing uniformization and specialization, and not towards the exceptional specimen representing evolution: the type that is originally very ill-suited to that environment, but carries a powerful survival and evolutionary potential. They cleverly invoke into their theories "selection", a verb, as an acting subject, a cognitive and decisive agent, a "deus ex machina", "who" is "working on the genetic variation that already is present in any new generation". Their kind of "Selector" would be in fact contraindicated for all those types, that were and are representing the "arrival of the fittest", those that were and are the spearheads of the evolution of their species.
The Neo-Darwinist theory coincides with and supports fabulously the favourite social worldview, because they open the way to attribute all human shortcomings, failures and incompetence to acausal occurrences, beyond the control and thus responsibility of any subject. It also gives the licence to manipulate selection towards "socially desired types". In an acausal and haphazard universe of ad hoc processes and "random mutations", also "selection" must be indiscriminate in order to be consequent to the inconsequences, producing even more confusion, instead of order. Doesn't anybody perceive, that the whole slapdash idea of materialism and acausality is violently contradicted by simple, quotidian reality? Or is everybody, with the exception of the thinking mini-minority, prevented to perceive it with all the possible instruments of brainwashing, in which science plays a key role by prostituting itself?
The crux of the trouble within the highly specialized and compartmentalized science is, that theories and arguments are based on minusculous particulars, not on holistic studies. Each specialist, being limited by his specialty and in the scope of his inquiry beyond it, by the frame given to him as facts, is essentially a reductionist, and sees and argues his subject as a closed system. Evidently, antitheses can be presented only within the limits that have already confined the theses. Thus the whole evolution of life on earth is being discussed on the level of the protozoon and around the question if a flagella attached to it could have developed by chance or only through "intelligent design"; its outcome deciding whether we live in an irrational or rational universe.
I have yet to meet a cock-sure neo-Darwinist able to approach rationally, as a probabilistic occurrence, that "there are some kinds of orchids as I read by Sir Alistair Hardy with flowers, which mimic, in colour, shape and smell the female form of certain insects and so offer sexual attraction to the males of these insect species; the excited spouses who come for the creative act, unwittingly, by carrying pollen, complete, instead, the sexual process for the flower." In this case the formation of the shape, the colour and the smell must have been simultaneous, neither of them having any value by itself; and all that to a specific model, which is directed not towards the present survival of the particular entity, but towards the future propagation of its kind.
An éclatant example for highly specialized and compartmentalized science basing its arguments and theories of "intelligent design" as evolutionary causation on minusculous particulars, and not on holistic studies, is the looking at such an enormous and all-involving subject as evolution through the super-microscopes of microbiology and on one of its findings. But even so, this theory would win playing against the neo-Darwinian random irrationalities, should its advocates object to the identification of their "intelligent designer" with the extraneous anthropomorphic deity of Moses' Genesis, or with the deity of any other instituted religion or belief-system.
Physiological intelligence, the intelligent reaction of organs tissues and cells to organismic demands and stresses, has been known for over half a century; as has been known, that even bacteria react fast with additional genetic immunity to new antibiotics. The by now through observation firmly established processes of genetic alterations at cellular levels as intelligent responses to changing environmental conditions in multicelled creatures, is translated by many microbiologists as "DNA reorganizes itself intelligently when organisms are environmentally stressed", and that "genomes respond to stress in many different species, from microbes to plants and animals, with the changes then passed on to succeeding generations". While this removes the "probabilistic ad hoc mutations", and replaces them with "intelligent chromosomal self-reorganizations as responses to stresses on organismic level", as the explanation of genomic changes, it still leaves the intelligent causation of the biological evolution of organisms at the chromosomal level, and attributes species diversity and evolution, and the harmony within ecological communities, to chromosomal intelligence.
According to holistic cosmovision, evolution is an open system, and happens on the level of every holon of the Natural Order, where all are the subjects, the instruments and the originators of Creation; each holon, in a holon-chain of continuous interactions (e.g. cell tissue organ organ-system organism ecological community planet) remaking itself in the image of its own values at its own level of existence, which come out of its own perception of its part as an individual and as a constituent through a chain of higher orders of the whole.
Another variation, which is a more rational speculation because it also includes its proposed reason for the phenomenon of an "evolutionary jump", but still leaving the sensing and the decision-making on chromosomal level, might be illustrated with W. Garstang's "Recapitulation Theory" and Arthur Koestler's thoughts and variations on it, which I will discuss in the following, mostly as foundations for further thoughts.
This theory is based on observational data on evolution; on the evolutionary lineage exhibiting a zigzag course in which periods of slow advance of constant direction appear to be separated by periods of very rapid remoulding towards a new direction; and on the presumption, that evolutionary changes are necessarily forced out by some environmental pressure, or dead end. Taking this conjecture as the motive, it starts right away with working out the possible biological mechanism through which an "evolutionary jump" may come about. It ignores, however, that in all the cases only a fraction of a species community responded to the "pressure", the rest continuing untouched on their accustomed-to path of low evolutionary gradient, without reaching a dead-end or extinction.
According to the theory, an ordinary jump has three physical stages: (1) undoing the existing relaxed and balanced state of rest or uniform motion by drawing back, taking position, flexing muscles, etc. This is a disequilibrated state, which has already nothing common with the original state of rest; (2) the jump proper, the efficiency and result of which depend very much on the extent and efficiency of the undoing and flexing energy-build-up of the first stage; (3) regaining equilibrium and consolidating a new state of rest or uniform motion.
In this form the analogy is applied to evolution, where the proper jump represents the evolutionary jump of a species, with the radical alteration of their mental and biological habits with the corresponding morphological and physiological changes, and the alterations in their relative environment. All this happens within a comparatively short time, resulting in increased evolutionary gradient, and requiring high energy-output, exhausting the system, which, needing rest and the regaining of equilibrium, enters into the third stage, where the achievements are stabilized, consolidated, concretized and perpetuated through the procreative mechanism and conditioning during the time of infancy. This is a period during which the previously described low gradient evolutionary movements with rules adjusted to the new life-style continue until the next jump is forced out by the environment.
According to that theory, as a stabilized and consolidated internal and external condition does not invite or provoke an evolutionary "jump" producing a new species, stability and consolidated conditions can be undone only by altered conditions in the environment. This is then reacted to by a flexible, potential- and energy-rich condition, suited to make a new evolutionary break-through. This condition exists in all organisms during the period of their development, that is their foetal and infantile stages.
Figure 14-3 shows Garstang's representation of his theory on the progress of evolution by paedogenesis. In it Z to Z9 is the progression of the fertilized eggs (zygotes), and A to A9 the resulting corresponding adult forms. The lines Zx to Ax are the ontogenic lines the transformation of eggs into adults , A to A9 the phylogenic one the evolution to higher forms. This latter represents, however, not a direct evolutionary progress, which would mean gerontogenesis, the development through fully adult structures. Evolution, in Garstang's theory, branches off at an early developmental (embryonic) stage from the ontogenic lines, and "retreats" back to the next zygote, and to a new departure. The result is a zigzag pattern (emphasized on the diagram), a forward-backward movement that Koestler characterized as "reculer pour mieux sauter" "drawing back to leap better".
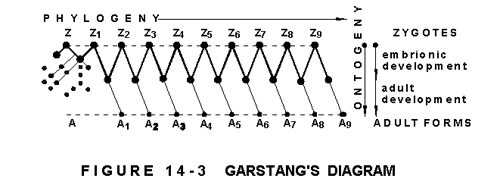
Garstang represents ontogenic development (which includes growth and consolidation) as a straight line (Zx to Ax), always starting from a new zygote (Zx), and ending in an adult form (Ax), that means simultaneously a dead end. It jumps the phylogenic ladder (towards higher forms) in the form of retreats from embryonic stages into higher zygotes. The jump is in fact a draw-back from where recommences a new, constant, linear development towards a new dead end, from which it is saved in time by a genetic mutation causing organismic changes as from the embryonic stage.
The most misleading or rather wrongly conceived are the expressions "draw back", "retrace steps", "reverse" and especially "regress", used in connection with reaching from the third stage of an existing state to the first stage of the new "jump", because an organism cannot regress from consolidation into new development, as man cannot regress from senescence into youth. He can stay young, however, which is the function of the mind; where young means flexible, dynamic, developable and developing. This is paedomorphosis, the process in evolution whereby adult forms retain young characteristics. Retain and not regress to it.
The diagram implies that the zygote, the fertilized egg that is actually an unicellular organism, genetically complete, capable to reproduce itself by cleavage, and produce, through further divisions, and according to the genetic blue-print (chromosomes) contained in its nucleus, a complete multi cellular organism (animal) is manifestly different at each rang of the evolutionary ladder, and that the life-cycle of each member of a group runs from Zx to Ax until the jump (or rather the retreat) comes, from which time on it will run from Zx+1 to A x+1.
In my view, "dead-end" is a relative concept and a subjective definition of a state of being. I will not enter into argument with one of our ancestors, the see-cucumber that is still there without nervous system and all which one of us is better off as far as plain survival is concerned, and who is more in a cul-de-sac at the moment. He is satisfied, for his billion years old state matches his billion years stagnant intellect. (The trouble would start if the cucumbers would "claim" like cabbages do their "rights to equality" on account of being relatives.) It is doubtful that after the innumerable splits into more and more species in evolutionary history, the "others" who have "stayed behind" have ever felt jealous, deceived or unhappy, or even inferior. They have stayed in their familiar, accustomed-to ecology niches, corresponding to their level of intellect, and can see no "dead-end" in it. The process is always more difficult and painful for the ones who "jump", for the spearheads of evolution.
Neither do I agree with the idea that dead-ends subjective concepts tick off automatically new break-throughs through biological changes. A dead-end, accordingly, would be characterized by some chemical, thermal, atmospheric and radiant energy conditions, producing certain types of protein molecules, which would catalyze some chemical reactions not anywhere, mind you, but inside the nucleus of the zygote, the fertilized egg of the organism in dead-end-trouble which, in turn, would produce a random or planned mutation, and off rides the new species or at least one specimen of it and evolution is once more saved from the cruel fate of dead-end-itis (but not from the cruel hands of the scientific Zeitgeist).
Inspired by the great many half-baked theories of that very scientific Zeitgeist, I propose a question that forever puzzled me: who and why for these double negativists, random variationists, chance mutationists, statistical transformationists, general ad-hoc-ists and mechano-biologists is being frustrated in its intension to regress towards protoplasmic existence by whom? who finds himself in a blind alley, and tries aimlessly to escape it, and why? who and why has programmed instinctive behaviour? who is that personified nature who has constructed that whom so, and why? who devised the mechanism for survival for whom? who and for what purpose wants to perpetuate himself and his species, or wants to evolve? and above all who and why wants to survive anyway, if there is no consciousness, intelligent mind and logical reason within each of those who-s and why-s, and behind the whole? And if there is, what are all the cock-eyed theories of mindless chance-happenings to conscious-less chance-occurrences about? Instead of standing in awe and admiration before the wonder of it all and trying to understand it!
According to the spirit of my thesis and while we are talking in metaphors : evolution is not the escape from blind alleys, but the answer to the call of the open spaces, of the wide horizons.
Based on my theory, I propose a different diagrammatic representation about the evolutionary process (see figure 14-4), commencing with the formation of the simplest biological systems. While this involves several steps, I denominate it as origin, and follow directly with the previously referred-to macro-molecules, of which the RNA and DNA are the ones forming the chromosomes, the genetic material of the cell, but which exist also in isolation as viruses.

The diagram shows the gradational series of the evolutionary "jumps" in direct line from the origin to Homo sapiens. Above the line are marked, where applicable, the biological and embryological definitions of the corresponding stages, and also, where known or suspected, the connecting links, the ones who jumped. Below the line are the biological classifications of those groups which contain the immediate others who stayed behind. The inclined lines represent their consolidation and conservation. Their slight forward inclination indicates a continuous, low-intensity evolution. The adult forms find themselves on these lines, their distance from the break indicating their grade of consolidation. The arrow points towards further consolidation, but does not represent a dead end with the exception of Homo, the link between the hominid apes and Homo sapiens. As evolution increases from left to right, the state of consolidation increases from the top downwards, and the further down a species finds itself on that line, the higher will be its relative persistence versus change, its racial inertia against evolution.
Each animal's or rather each and every living organism's ontogeny starts at the origin, and shares the evolutionary straight until it reaches the point of the branching off characteristic to its species, from where it continues quasi vertically, repeating its own remaining evolutionary path until its (and its species') particular point of evolutionary stage and consolidation, thus following its phylogenic line. Other species could also be included into the diagram to demonstrate relative evolutionary tendencies. These would branch off at their specific point, with the evolutionary gradients of the jump inclined in proportion to their relation to maximum achievement, but would never cross the consolidation vector of the next higher order, accommodating themselves between the limits of their evolutionary potentials defined by their own moves and those of the others.
Any consolidated developmental state is suitable only for the condition for which it was fashioned. However, this condition is being challenged constantly by the other species' reactions to it, and by the similar kinds of developments of all the other species constituting that particular ecological community metabolism. The evolutionary steps, with their ensuing widening horizons are accumulative, and are the functions and the tracer stimuli of the particular intellect. Evidently, intellectual differentiation is followed by an evolutionary split, and by accelerated and decelerated evolutionary tendencies. (That some reptiles could develop cerebral cortex, and open the way for further extensive evolution as mammals, a group of them had proven back in the Triassic age, some 200 million years ago; that the "ones who have stayed behind" haven't got the stuff for it, they are proving ever since.)
The new relative environment, which also represents a new ecology-niche within the ecological community, is by the very nature of the evolutionary change much more complex, and places much higher demands on each individual. These demands have to be met individually. The ones who fail, face the problem of no return, and the fate of the relatively unfit and incompetent. Judging by the proportions of the progressives and the others who stayed behind in evolutionary history e.g. mammal-like reptiles against reptiles in the late Triassic, or first primates against the rest of the mammals in the early Eocene the relative numbers of the individuals forming the new species is, in the epoch of the separation, insignificant. Looking at it from another angle, there is nothing derogatory in staying behind, as long as each individual keeps and fulfils with dignity the place in the system that corresponds with its own intellectual potentials and willingness, for that system created and filled all ecology-niches, and produced a well-balanced order called planetary life, the dynamic equilibrium of which was endangered only when some species made a somatotonic revolution, and forced their physical presence well above their mental capacity.
Where the individuals of a species are aggregated into social groups, and carry out most of their life-functions like obtaining food, shelter and protection specialised or not, but together, social systems are developing, "pecking-rights" established according to individual merits, and the leaders become responsible and take the decisions for the group. These decisions establish the ecology-niche, the relative environment and the culture in accordance with the majority's capacity and not its decision! , which majority occupies the central half of the intelligence range of the group, and represent about 86% of its numbers (see the standard deviation curve as explained in connection with figure 14-2). The 7% on the lower end (between -3 and -1,5 SD) are intellectually incapable to survive under the conditions provided by the relative environment as created by the group, and are falling out somewhere between birth and maturity, being, however, constantly replaced by the over-breeding of the lower elements of the central half.
From a biological point of view two main factors were the causes and the consequences of relatively higher evolutionary gradients: a higher degree of cephalization, and a generalized structure that confers a greater degree of functional plasticity. Both are tools and imply an elastic-dynamic, non-mechanical and non-calcified state, typical to the embryonic and pre-sexual (infantile) stages of development. These are, however, also the stages in which the mechanizing and calcifying processes are the most active both on organismic and environmental levels. These processes would have the effect that by the time the individual reaches maturity, all its manifestations are hardened into mechanical actions should this be nest-building, or social, philosophical and religious convictions and behaviour which it will transmit to further generations through genetic (sexual reproduction) and social (conditioning) channels. All are relatively degenerating, because stability, which is always stagnant, equals relative regression, for the rest is in movement and passes by. The necessity to move in order to keep up with the general evolution is called the "red queen effect", after the Queen of Hearts telling Alice (of Wonderland fame), that in her kingdom everyone must run as fast as can, just to stay in one place.
In biology, neoteny is the retention of embryonic or pre-sexual conditions in some organs, although the animal becomes otherwise sexually mature. Sexual maturity defines in this context not the fitness to copulate with a member of the opposite sex (or, following the present fad, of the proper one), but the fitness to transmit through the pro-creative mechanism real and representative acquired psycho-physical characteristics, and thus consolidate the new characteristics in the new species. The emphasis should be, however, not on sexual but on mental maturity: the stage of development when the individual has both the conditions and the intellectual capacity to use the potentialities that embryonic and pre-sexual conditions offer mental and physical elasticity and plasticity and remould itself accordingly. The change marked unto the chromosomes is then genetically transmitted, which is where the sexual maturity enters. Logically, neoteny only has value in progressive evolutionary development, if there is an agent that can make use of the given potentialities. This agent is the mind. On the other hand, adding mental to sexual maturity, neoteny becomes a condition quasi sine qua non for evolutionary progress. And here enters the peaceful Avatar into biology: "unless you will become like little children . ."; like little children: elastic, plastic and dynamic in mind and body. This is paedogenesis, but not evolution (personified) randomly retracing its steps to an earlier, more primitive stage, and making a try for a fresh start by introducing, through a lucky chance-mutation, some also random, "useful evolutionary novelties" in the larval or embryonic stage of the ancestor.
I want to emphasize a crucial point: mental development is not the consequence of chromosomal mutation: chromosomal mutation is the recording of the mental development on the genetic blueprint. The first amphibians did not come out of the waters because they grew lungs by some weird genetic chance, and were forced by it to do so, or because they got stranded in a puddle and were waiting there patiently for the same weird genetic chance to save them before the puddle dries out: they wanted to go unto the dry land because it was fascinating "because it was there!" , ergo their organism has produced the lungs and the legs, and all the morphological, histological and physiological changes necessary to serve them in that intellectual adventure. The initiative was theirs: the mental force of the being, directed towards an aim, challenged and energized its organism on every integrarchic level to respond positively to an evolutionary demand, and using what is called its physiological intelligence, develop a new physiological technology, and reconstruct itself according to the best of the actual intellectual abilities on every level of its being. But there is yet another crucial point: the amphibians or any organism for that matter did not adapt themselves to the physical environment that they encountered, but created for themselves a new particular mental environment incorporating the physical one the way they conceived it, and moulded their own bodily structure to their own reality. Naturally there enters trial and error into this process, but that of the creative designer, and not that of the passive gambler.
The mental composition of an organism holon has been expressed (equation 3) and discussed earlier in this book, as has been the complex mechanism of the Integrarchic System of the Natural Order. All apply to what have been exposed above, which would need no further elaboration except for a most important fact that must be stressed once more: species do not evolve, individuals do. Individuals have concrete existence; species is a biological classification. If many individuals possess quasi identical characteristics, they are called a species. The high degree of similarity is, no doubt, the result of having the same ancestry, of inter-breeding, and of sharing the same functions within their ecological communities, but this does not account for a collective evolution, neither does it make a social concept out of the whole species. Species community refers only to those of the same species occupying the same ecology niche witrhin an ecological community.
Individuals of the same species forming close social units can undergo genetic drift expressed in intellectual and character differences, and differentiate into two sub-species, first psychologically, then morphologically alienated. Finally, the splitting consists of individual actions in collective manifestation, as are the psychological and morphological adaptations to the newly created situation. Nevertheless, individual evolutionary process can only be perpetuated (or wiped out) through the interrelationship of individuals, who independently and in unity perform the actions characteristic to their life-expression, and conceptualized by their collective.
I have to interject here that there exist "non-local" connections and orchestrations on another dimension between species- and ecological communities and their evolutions, as influential in the process as are the physical ones, but not having general acceptance by far, I do not involve them in this thesis. Scientists, regardless how knowledgeable, will never recognize this, unless they are at the same time also of a creative spirit.
The process, that is the fundamental mechanism of evolutionary diversification, will be further elaborated on after developing the evolutionary model (figure 14-5), starting from the beginning.
When the earth has formed itself about five billion years Before Now it was a whirling interstellar gas. Cooling down it liquefied, then gained a solid crust, the lithosphere, and an atmosphere. The latter consisted of hydrogen combining with carbon, with nitrogen or with oxygen, forming methane (CH4), ammonia (NH3) and water (H2O) respectively. With further cooling the water condensed, forming rivers and seas.
Looking at a planet with this kind of an atmosphere, we would classify it now as unsuitable to support life. Yet exactly these compounds have formed themselves absorbing solar energy into more complex molecules, aminoacids, then carbohydrates, lipids, proteins viruses, bacteria, cells what we define as living organisms. Thus a biosphere has developed, involving more and more the lithosphere and the atmosphere, transforming more and more of them into itself. In this biological process the methane-ammonia atmosphere was changed into a carbondioxide-nitrogen one. A further sub-cellular development produced the process called photosynthesis, which, with an efficient use of light-energy, consumes carbondioxide to build up complex molecules, and liberates oxygen. Anaerobic respiration changed thus into the much more efficient aerobic one. It has to be pondered upon that these microscopic creatures (for no other existed), all living in the waters, have changed twice and completely the whole atmosphere of the planet improving themselves constantly with it before the first multi-cellular organism could ever even start to develop. They took their time, no doubt: this period lasted from three and a half to one billion years Before Now.
What has evolved primarily was the Planet's Life as a whole, in a protoplasmic continuity, developing more and more complex compounds, using the basic elements of solar energy either directly (like in plant cells), expanding into innumerable varieties through stratifications, subdivisions, differentiations and refinements, increasing all the time their complexity, each fraction forming a quasi-self-contained open system a holon continuously elaborating its own development, while exercising its own logic and creativity; each as a symbiotic aspect in the evolving pattern of that Life, all in accordance with a Higher Ordering Principle.
According to my thesis, the particular intellect that enters in every form of life-manifestation, organic or inorganic, as an ubiquitous constituent of the Psycho-Physical Reality, is directly proportional to the evolutionary level of an organism. This was already implied in connection with the evolutionary diagram (figure 14-4), where intellect with evolution or evolution with intellect grows from the origin towards the right, into the far off horizons.
As evolution has (to my knowledge) no factor, but intelligence mind in its qualitative manifestation has, I am using the latter to define evolutionary progress, and calibrate the horizontal co-ordinate accordingly. The progress being as discussed earlier exponential, the calibration is logarithmic, the Origin being marked by the unity, and the actual median intelligence factor or evolutionary factor of Homo sapiens by 100. This marking is arbitrary, but as the scale expresses relationships and not absolute values, it is as valid as any other figure would be.
On the vertical co-ordinate I am introducing another dimensionless property: inertia. This relative persistence includes the conservatism of from morphological elements to mental habits, and is defined as the passive resistance to change, manifested by a holon (animate or inanimate) as function of its own state of energy and stimulus-response time. It is intrinsic in the holon as it is in the Universe on all time-bound planes. Stimulus-response time refers here to rational mentation and not to automatic reflexes. Particular inertia influences and is influenced by the momentary inertial state of the Universe.
As inertia can be related to the time it takes for a certain cause to take a certain effect on a certain object, the resistance to change can be defined by the time it took for a certain development to realize itself. Phylogeny, the evolutionary history of organisms provides exactly the right scale, which, to represent the correct proportions, is once more logarithmic and starts with the year 15.109 BN. For obvious reasons I have tempered with the scale, defining the mark 102 as present, and reversing at this point the regress into the past to the progress into the future in the same logarithmic scale.
The diagram is based on much intuition as it is on hard facts, and, admitting that intuitions can be sometimes as erroneous as can be hard facts, it is open for discussion and improvement.
The maximum evolutionary gradient, marking the gradational series of the highest evolutionary progresses, connecting the highest evolutionary forms respective to the epoch, in direct line of descent ending in modern man, is inclined 45o, suggesting that the maximum exerted evolutionary force has equalled the relative persistence. It also marks the line of relative maximum achievement due to relative maximum effort. The non-achievement line the path of least action is vertical in the diagram, and sets the other limit to evolution. Evidently, negative gradient represents regression.
The phylogeny of a species marked by a single line would imply complete homogeneity within that species in every sense, that would ab ovo prevent diversification. More true to reality will be if the probable areas they have occupied and are occupying in the evolutionary/intellectual scale will be shown, neglecting aberrant development (except for Homo sapiens) and extinct branches, and remembering that there are no re-crossings of particular evolutionary paths after the splits have been established.
Perceiving the Universe as a dynamic process, where logic, consistency and aesthetics are basic principles, I believe that a diagram (or a mathematical equation), the closer its gets to the reality of the processes it want to represent, the more it will exhibit these elegant qualities. This would firstly mean that there can be no abrupt break at the point of a split, but a curve, to which the evolutionary gradient is tangential at the point marking the change. Considering further that the ontogenic cycle of every organism starts with the aggregation of the basic elements that is at the origin of organic life its development follows the evolutionary gradient of maximum effort, until it reaches its respective point of departure, from where it will describe a trajectory, which I suggest is a parabola (ballistic curve, if you wish), terminating at the present evolutionary state. This trajectory that is the path of least effort is the function of the attitude of the evolutionary gradient and of the energy-conditions of the organism at the point of departure. As evolution has accelerated steadily during its history, the ontogenic path of an organism can be compared to that of a ball starting at the origin of the slope, gaining speed until the point of departure, then continuing according to its own inertial conditions, being deflected by the 'gravity' of relative persistence acting in the direction of the future and the 'atmospheric conditions' of the evolutionary environment.
Remember, the gradient is a concretum: it is the resultant of two vectors: evolutionary force (active mind) and relative persistence (inertia). The energy-condition of the organism at the point of departure is proportional to the square of its evolutionary factor, that this being a study of relationships defines its relative ontogenic path.
Considering well the above presented evolutionary diagram, and also the fact that the Universe is a living and evolving entity, where diversification and individual and through it species evolution is the natural trend, an evolutionary split represents in fact not a jump of those who are continuing ahead smoothly on their evolutionary path, but the deflection of those who cannot keep up with it. The new general circumstances demand higher mental faculties, and throw-backs will fail and fall out through self-elimination, causing the fast extinction of the connecting population; one of the possible reasons why 'missing links' are so hard to find between species of higher evolutionary states.
There are two alternative causes that can change the parabolic trajectory of a deflected species: re-applied evolutionary force, and acquired additional inertia. Both result in an evolutionary split. The first one produces a secondary gradient tangentially to the curved path at the point of the split, marking a new intergrading species. Evidently, the secondary evolutionary gradient will be less inclined than the primary one, and so will be lesser and lesser the gradients of later evolutionary progresses resulting in further differentiations. Thus the ontogenic path of an organism can consist of several straight paths representing forced evolutionary progress and several curved ones of inertial persistence, all in an unbroken continuity. It also appears to be a rule and a logical inference, that new evolutionary efforts can only be initiated from the actual evolutionary state, and never from a preceding one, which partly accounts for the fact that evolutionary paths do not cross. The lower the order of the evolutionary gradients the closer they get to the path of least action the less is the evolutionary/intellect pressure, and the less is the differentiation.
The acquired additional inertia corresponds with a state where the temporary environmental conditions in the actual ecology niche of the species become so easy (e.g. by exploiting and living it up), that the majority by not using it loses its evolutionary potential that is basically its intellect and becomes relatively or absolutely regressive, its evolutionary path deflecting even more towards the path of no achievement. The rest, the ones on the upper end of the intellect spectrum of their species, continue on their higher evolutionary achievement path, the others' digression of which in the negative direction creates the split. Those who regress even further get extinct, because they fall back into another ecology niche in which they haven't got the capacity to survive. This is why I have shown on the diagram the path of least effort as the one which marks the minimum long term survival chances for the corresponding species.
The space between the co-ordinate axis of time the vertical marked with unitary intellect and the primary evolutionary gradient to the point of the origin of organic life, with the to this point corresponding trajectory, represents the evolution of the inorganic world. It marks the transformation and building up of primordial energy through mind into subatomic particles, the hydrogen atom, all the elements according to the periodic law (Mendeleyev), and their combinations into compounds of growing complexity, all in accordance with the thesis exposed in a previous chapter and expressed in equation 3.
On the primary evolutionary gradient are marked the appearances of the actually highest evolved organisms, which represent at the same time the direct lineage of ancestry of Homo sapiens. Very important characteristic of this linage is that it clearly exhibits anamorphosis, an organism's exercise of its potentialities of evolution to create an improved, new form of life, driven expressly by intellect-pressure, and not by some extinction menacing environmental occurrence. For instance, the first peaceful little mammal-like reptile becoming warm-blooded, placental and starting to grow a bit of hair appeared not when the reptiles have got themselves into a blind alley, or faced the danger of extinction: on the very contrary, they appeared at the time of the reptilian outburst, when to use a favourite human expression they dominated the world. The other reptiles were comfortable, well adapted and in the pink, but he took on the task to improve himself, grew the neophalium (cerebral cortex), a new extension to his existing brain, and founded on the strength of it the mammalian class. The same happened about 120 million years later, when his descendants were in full swing, and the primate order was originated by a peaceful, small tree shrew type ancestor of ours. The story repeats itself from the beginning right up to the present kind of Homo sapiens, and so it will go on further. Out of any group, those who have finally succeeded, prevailed and went on forming the superior lines, were the ones with bigger brains, and not the ones with the big bones and muscles, or with the big guts. What they all had in common on the entire line of evolution was mental over-intensity, overly fast reactions, non-aggressiveness, quietude and restraint, physical agility and youthful manner of appearance, and all were fragile, exposed and unprotected in structure relative to the prevailing qualities of their respective groups. On the other hand, those who tried to solve the problem of mere survival with bones and muscles (somatic or mesodermic structures) aggression grew first to tremendous numbers and sizes, then suddenly got extinct.
There was and is also a third kind, resorting to the third alternative: his guts (visceric or endodermic structures). It is decidedly regressive. While the other two use their brains and their muscles respectively to change their environment the first evolving, the second subduing it the third one attaches and adjusts himself completely to it for pure survival, tending towards simplification, becoming thus, to a lesser or higher degree, harmlessly or harmfully, parasitic. While they are found mostly between viruses, germs, the protozoa and some lower metazoa, they appear even at the highest evolutionary level as aberrations within a species, and belong to the fall-outs at evolutionary changes.
Whatever line of thought is followed, the conclusion is that evolution is a cerebral first principle and not a somatic or visceral process. The corresponding organs as described earlier have developed out of the three (there are no more) germinal layers: the ectoderm, mesoderm and endoderm. Both the endoderm and the mesoderm, and the from these developed visceral and somatic sub-systems were results of the courses of action taken by the mind the first principle through its instrument, the cerebrum, according to its perception of the demands of its own development and of its environment, the correlation of its impressions, the construction of systems of thought, and the setting out of these courses of action. Evidently, these auxiliary systems gained autonomy through differentiated mental functions, the progress of the individual organisms depending further on the correct equilibrium between the mind in command, and the delegated brain centres the autonomous or vegetative nervous system in charge of the sub-systems, remembering well that in the tripartite division the mind is the primordial element, and that the delegated centres and further sub-centres are of a lower order, in level with the instincts, but can consequently become just as domineering as instincts, fixed pattern of mental or physical behaviours, can. With increasing intellect increased also the differentiation, but the morphological tri-polarity, like the three germinal layers, and as the expression of the tonic tri-polarity of the psyche, perpetuated, only the variations became more imaginative and vivid on the theme composed of the three basic notes.
The peaceful, rational mind is the underlying principle of the Universe, and thus of every fraction of the living Earth. The "law of the jungle", glorifying aggression and devastation, is a human invention, not that of the jungle. It is the credo of those who can see in the symbiotic evolution of a planetary life-community only its catabolic phase, that even in this action is not destructive, but organized and selective, promoting the evolution of itself through the prevalence of its most vigorous and evolving constituents.
Eternal is The Ground, the Mind, the cause and mover, the underlying principle of all existence. The Mind that causes Chaos coalesce into Form and Action.
Whatever exists in material form as an integral part of the Universe, is the manifestation of the Mind, that permeates and sustains the All, and causes the flux and reflux, evolution and involution, synthesis and analysis, condensation and dissipation; breaking into infinite harmonies of the heartbeat of a galaxy, the heartbeat of a planet, the heartbeat of a mountain, the heartbeat of an atom, and the heartbeat of a man.
As the all-transcendent Ground of all existence is changeless, static, pure Being, so is Its manifestation of incessant change, dynamic Becoming, continuous Creation, in accord with the basic note and its infinite harmonics, the music of the spheres. Thought and action, planning and execution are the rhythms of becoming. Every healthy fraction of the universe possessing the individual spark, has the drive to become, to grow, to progress, to evolve, to know, to create. The expressions of the will to live are inherent in each and every holon of each and every stratum of the Natural Order. Being integral part of the Universe, the fraction is one with the Infinite Mind.
Harmony is a foundation principle of the Universe. It is manifested in love, in the strong and weak forces, the electric, magnetic and gravitational interactions, the chemical valence forces, cohesion, adhesion, right up to symbiosis and synergy, the harmony of the music of the spheres, all that moves, sustains and "holds bound into one volume all the leaves whose flight is scattered through the universe around" (Dante).
Matter, the physical manifestation of atoms and galaxies, is time-dependent and finite: its existence in a certain particular form or complex is limited by two events: its coalescing and its disintegration as such. If the second event does not occur, the body will continue existing in that same form indefinitely, without loosing its potentiality to change, or its quality of being finite. An element, like hydrogen, carbon, iron, that had its origin in time through nuclear fusion (according to the fixed rules and flexible strategies of its being), or any stable combination of these elements, that originated in chemical reactions, can, if isolated as a closed system, continue to maintain that form indefinitely. An organic cell, the common unit of structure of all organic existence, this highly complex, multi-levelled organization of elements, can, if alimented, kept under suitable environmental conditions and protected from its predators, keep on living and multiplying itself through division ad infinitum, meaning potential immortality. Except for one fundamental quality: existing on the psycho-physical plane, the basic principle of which is change, development, becoming, the will to live of every particle of the universe expresses itself in the will to grow, to progress, to evolve, to know, to create; transcending the comfortable, eventless deathlessness. Because of this, inanimate and animate matter chooses into concentration instead of dispersion, combine into complex compounds of higher order instead of decompose into a lower one, increase syntropy instead of entropy, and provide variation and evolution by creating new forms and consolidating it through sexual reproduction; all containing the element of free will, and all representing a finite existence of expression within one form.
The life-cycles of the different life-forms vary greatly between a fraction of a second and billions of years: in an adult human, more than quarter of a million cells die and just as many are born within every second. We know nothing of the "tragedy". Neither is it one. For the essence, that is the cell, remains and continues creating, sustaining and representing life in the form he has developed itself to do so.
For every particle of the universe lives, and all physically manifested life consists of a Mind and a Body, representing a fraction of the Sum of Being. And so does man. And considering man, all universe must be considered, because a thesis and this is of fundamental importance can be applied to humans only if it can be applied to each and every fraction of the Natural Order.
The body is mortal, for it has a beginning, an existence and an end. It is mortal out of the logic of the very Principle it incarnates, and for the very reason that it is brought into existence by that same Principle: it is Its vehicle of expression and growth. Not as an isolated personal expression or personal growth, but those of the whole Principle.
Beginning and end, birth and death, are not the limiting marks of an isolated existence, but mile-stones of an infinite world-line, where the end of one kind of existence marks the beginning of another kind, and death in one sphere of life the birth in another. It is the law of change, where the variation of the infinite rhythm is marked by differentiation into continuous rebirths: the evolution of a Living Universe.
Imre von Soos © 1984, 1987, 1995, 2001, 2004, 2006
|