|
TRANSLATE THIS ARTICLE
Integral World: Exploring Theories of Everything
An independent forum for a critical discussion of the integral philosophy of Ken Wilber
 Andrew P. Smith, who has a background in molecular biology, neuroscience and pharmacology, is author of e-books Worlds within Worlds and the novel Noosphere II, which are both available online. He has recently self-published "The Dimensions of Experience: A Natural History of Consciousness" (Xlibris, 2008). Andrew P. Smith, who has a background in molecular biology, neuroscience and pharmacology, is author of e-books Worlds within Worlds and the novel Noosphere II, which are both available online. He has recently self-published "The Dimensions of Experience: A Natural History of Consciousness" (Xlibris, 2008). SEE MORE ESSAYS WRITTEN BY ANDY SMITH How History
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A) Dimensions of the Physical Level | ||
|---|---|---|
| Dimensions | Representative stage | Examples |
| 0 | atoms | hydrogen, helium, oxygen |
| 1 | small molecules | amino acids, sugars |
| 2 | simple biological polymers | peptides, RNA, cellulose |
| 3 | complex biological polymers | proteins, DNA |
| 4 | active macromolecules | receptors, enzymes |
| 5 | macromolecular networks | citric acid cycle; electron transport chain |
| B) Dimensions of the Biological Level | ||
| Dimensions | Representative stage | Examples |
| 0 | emerging cells | photosynthetic bacteria and algae |
| 1 | primitive cell colonies | archaea colonies, bacteria colonies |
| 2 | advanced cell colonies | eukaryotic colonies, multicellular tissues |
| 3 | simple cell networks | motor unit, simple sensory system |
| 4 | complex cell networks | neuronal ganglia |
| 5 | small world cell networks | complex ganglia, brain |
| C) Dimensions of the Behavioral Level | ||
| Dimensions | Representative stage | Examples |
| 0 | simple organisms | Volvox; Fungi; sponges |
| 1 | simple colonies | coral reef, Hydrazoan colonies |
| 2 | differentiated colonies | crab colony |
| 3 | simple societies | ant colony; bee hive |
| 4 | family-based societies | bird and mammal societies |
| 5 | band-based societies | primate societies; human societies |
Hierarchical organization thus combines a few units or components at one level into a larger number of units or components at a higher level. Moreover, in nature, many hierarchies are also holarchies (see Box 2). In holarchies, a single unit or component at one level includes several, generally a large number of, units or components on the immediately lower level. An amino acid contains one hundred or more atoms. A peptide contains dozens or hundreds of amino acids. A complex protein can contain a dozen or more peptides. Such components are called holons, a term referring to the fact that they are simultaneously both wholes and parts (Koestler 1969). Thus an amino acid is a whole, composed of many atoms, but it's also a part, contained within a peptide. A peptide is composed of many amino acids, but in turn is contained within a protein.
The key feature of holons relevant to our present discussion is that their relationships with each other approximate those of dimensions. Consider an atom. In the scientific view, an atom is a three-dimensional form of existence. However, it is so very small, compared to the world with which we are ordinarily familiar, that we might regard it as approximating a point, a zero-dimensional body. From this vantage, small molecules such as amino acids, which are composed of one hundred or more atoms, are one-dimensional bodies. Likewise, peptides, which are composed of dozens or hundreds of amino acids, are one-dimensional relative to an individual amino acid, and two-dimensional relative to an atom. Similarly, complex proteins are three-dimensional with respect to atoms. Holons thus have a relationship to each other that I refer to as natural dimensions. They are not exactly like mathematical dimensions. The characteristic feature of mathematical dimensions is that they have a relationship of infinity to each other. A line, a one-dimensional figure, contains an infinite number of zero-dimensional points. A plane, a two-dimensional figure, contains an infinite number of lines. A three-dimensional cube or sphere contains an infinite number of planes.
Holarchical stages have a relationship to each other that is not infinity, but which does approach it; the relationship is of many to one. There are many atoms in what I call a one-dimensional molecule, but not an infinite number. There are many, but not an infinite number of, one-dimensional molecules in a two-dimensional molecule; there are many two-dimensional molecules in a three-dimensional molecule.
Another important difference between mathematical and natural dimensions is that in the latter, the relationship is not (necessarily) one of point to line to plane to cube or sphere. When I claim that certain molecules are one-dimensional with respect to their component atoms, I don't mean that their atoms are arranged like beads in a necklace (though sometimes in fact they are).[19] I mean that the atom, the zero-dimensional body, is the fundamental unit that composes the molecule. There is ordinarily no stable stage or state between the one and the other. In nature, as Francis Crick (1966) pointed out, we generally go from atoms to molecules of a certain minimum size.[20] Likewise, what I call a two-dimensional molecule does not necessarily have a planar appearance. It's two-dimensional because it contains a large group of one-dimensional molecules, which in turn contains a group of zero-dimensional atoms. And similarly with the third dimension.
We could therefore say that mathematical dimensions are an abstract ideal of what is approximated by actual dimensions as we find them in natural holarchies. Yet there is a close relationship between them, because as we will see, mathematical dimensions of a particular degree are strongly correlated with natural dimensions of the same degree. I have already hinted at this in the earlier discussion of the visual system, and many more examples will follow. So, for example, when I refer to a complex protein as a three dimensional molecule, I mean that it is three hierarchical stages higher than an atom, but also that it has features or properties that involve three dimensions of space.
So far I have considered only dimensions of space, but natural holarchies may also incorporate dimensions of time. Consider a complex protein, which I have provided as an example of a three-dimensional molecule. Many proteins exist not only in space, but also in time. That is to say, in order to understand their function, we not only have to take into account their spatial dimensions—their particular shape or conformation—but also that these dimensions can change during time.
For example, an enzyme molecule catalyzes the conversion of some substance, called a substrate (usually a smaller, one-dimensional molecule) to another substance (called the product) by changing its shape in a certain way. To understand the enzyme molecule, or to perceive its complete existence, we must therefore observe it not only in space, but also over a certain period of time. During this period of time, which is known as the enzyme's catalytic cycle, the enzyme molecule will typically change its conformation from one shape which allows it to interact with its substrate to a second, different shape that allows it to release its newly formed product. An enzyme molecule seen over this entire period of time is therefore a four-dimensional body with respect to its atoms. It has three spatial dimensions and one temporal dimension. This contrasts sharply with its component atoms or amino acids, most of which do not exist, in a functional sense, in these additional spatial and temporal dimensions.[21]
Temporal holarchies, like spatial ones, may follow Changizi's combinatorial rule, in which a small number of components at one level have the potential to combine into a larger number at a higher level. Each of the multiple peptides or subunits typically found in most enzymes may exist in two or more different conformations, which may combine to form multiple activity states (Monod et al. 1963; Koshland et al. 1966; Changeux 1998). Moreover, other activity states of an enzyme may exist depending on such factors as whether it interacts with certain small molecules (known as regulators), gains or loses a molecular group such as phosphate at a specific site, or interacts with still other enzymes (Chock and Stadtman 1977; Johnson and Barford 1993). Living cells use such processes in order to adapt a single enzyme to multiple purposes at different times, but the point here is that such regulation transforms a single three dimensional protein into a four dimensional process composed of many different temporal states of that protein.
The limits of holarchy.
To summarize the discussion so far, nature creates new forms of life by joining units, called holons, into more complex holons. Atoms form amino acids; amino acids form proteins; proteins form larger structures. Each new form of life may be considered higher or more complex than any of its individual holons. Again, without trying to define complexity precisely, I will just note that each new holon has emergent properties, ones not found in its component holons. Thus amino acids have properties not found in their individual atoms, peptides have properties not exhibited by individual amino acids, and so on. Examples of such properties will be provided later, when I discuss these forms of existence in more detail. We might imagine that new holons could be created endlessly in this manner, by simply combining holons at one stage or dimension into ever larger groups. In nature, however, this process does not, and cannot, go on forever. It reaches a definite limit.
Why should this happen? The organization of holons as I have described them so far is relatively simple. Each is composed of a great number of similar holons of a lower dimension. Because all of these lower-order holons are similar, and because they are organized in a relatively simple manner, there are limits to what they can accomplish. In particular, holons of this kind can't reproduce themselves. A molecule, for example, no matter how complex, generally can't on its own divide and produce two identical copies of itself.[22]
For reproduction to occur, a somewhat different kind of organization must emerge, one in which many different kind of holons are put together. In the evolution of molecules, this occurred with the emergence of the cell. Unlike an amino acid, which contains only atoms, or a protein, which contains only amino acids, a cell contains many different kinds of holons. In fact, it contains all the different kinds of molecular holons that are found in nature. In any cell, we find individual atoms; small molecules such as amino acids and nucleic acids; simple polymers such as proteins and DNA and RNA; and still more complex groupings of these holons.
The organization of a cell, on the one hand, and that of any molecule, however complex, on the other, represent two different kinds of hierarchy (see Box 2). As I noted earlier, holarchy is a special type of hierarchy in which members of higher stages include members of lower stages. Thus proteins include amino acids, and amino acids include atoms. Holarchy is exemplified by a series of Chinese boxes, each of which is nested within the next (see Fig. 1A). In addition to nested hierarchy or holarchy, however, hierarchy can also take a non-nested form, and this is characteristic of the cell. A cell is like a box which contains many separate boxes within itself (see Fig. 1B). Each of these boxes, itself, may be like a traditional Chinese box, with the one-within-the next form. So I refer to the cell's organization as mixed hierarchical, meaning it contains both nested (holarchical) and non-nested hierarchical structures. [23]
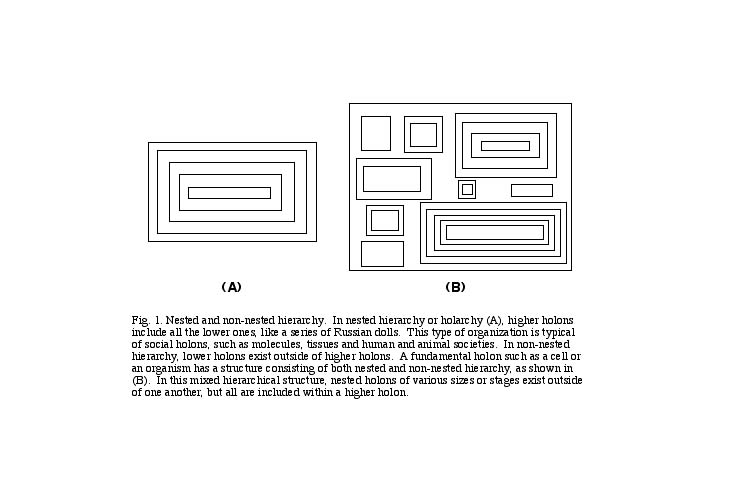
Cells therefore represent a profound transition point in nature, in which one level of existence, which I simply call the physical, is completed, and a new one, the biological, begins. For this reason, I call cells fundamental holons, fundamental to a new level of life. I also refer to them as individual or autonomous holons, since many types of cells are capable of existing on their own, outside of higher stages consisting of many cells.[24] It's their ability to reproduce that provides them with this autonomy. Atoms, though they do not reproduce, are also capable of existing outside of higher-order holons (carbon, oxygen, nitrogen, sodium and many other atoms can exist without being joined with other kinds of atoms into molecules), and so I also consider them as autonomous or fundamental. In contrast, holons such as molecules I refer to as social or intermediate holons, since they are composed of groups of autonomous holons, and form intermediate stages within a single level of existence. They generally cannot exist outside of higher-order holons. Thus individual protein or nucleic acid molecules, for example, are not found outside of cells or organisms, except in the artificial conditions of the laboratory.
Because a cell is not purely holarchical in its organization, we can't specify its relationship to its components in terms of natural dimensions. But since it begins a new level of existence, we can simply repeat the process we just went through with atoms and molecules. That is, we can begin by taking the cell as a zero-dimensional body or a point, just as we did with the atom before. While the cell is obviously not zero-dimensional with respect to its component atoms or molecules, it is zero-dimensional with respect to the new level which it is beginning. From this point of view, higher dimensional stages are exemplified by certain kinds of organizations of cells, including colonies of micro-organisms as well as various tissues within organisms (see Table 1B).
We can also identify multicellular holons that have an existence in time as well as in space, just as we saw was the case with certain kinds of molecules. A good example of this is found in the brain, where groups of highly connected neurons form networks. The neurons within such a network repeatedly send and receive impulses to and from one another, forming a pattern of activity that circulates in the network. The active network thus forms a four-dimensional holon, consisting of the three dimensional neural network over some period of time. And a particular network may have the potential to exist in multiple such states.
Again, however, we eventually reach a limit, beyond which pure holarchical organization does not pass. While individual cells can reproduce, multicellular organizations generally cannot. For example, a colony of bacteria can grow in size, through the reproduction of its individual members, but the colony itself can't reproduce as a single unit. Evolution solved this problem in basically the same way as it did when cells emerged: by creating a new form of life composed of many different kinds of holons, namely, the organism. Just as the cell contains all the different kinds of molecular holons, so the organism contains all the different kinds of multicellular holons. In an organism, we find individual cells, individual tissues of various kinds, and individual organs, all integrated into a single large holon. Like the cell, and unlike its component tissues and organs, an organism manifests the mixed hierarchical organization. The distinction of these two kinds of hierarchical organization in both cells and organisms has also been noted by Petersson (1996) and Valentine (2003).
Just as the cell completes the physical level and initiates the biological level, the organism completes the biological level and begins what I call the behavioral level. And just as we took first atoms, then cells, as zero-dimensional holons, so can we take organisms, understanding that they are zero-dimensional with respect to the level they are beginning. From this point of view, a one-dimensional body is a small group of related organisms, such as a colony. Still higher dimensional forms of organization are illustrated by more complex forms of social organization (see Table 1C).[25]
As with the physical and biological level stages, the stages on the behavioral level may exhibit dimensions of time as well as space. Organisms in complex societies interact with each other in certain patterns—behavioral displays, for example, and in the case of our own species, through language.
Evolution by socialization.
To summarize the discussion so far, life can be understood as evolving in the form of a holarchy, in which simpler forms of existence combine into more complex forms. Such combinations proceed through several fairly definable stages, until a transition point is reached in which an entirely new kind of organization evolves. The cell represents one such transition point, completing the physical level of existence and beginning a new level, the biological. The organism, analogously, completes the biological level and begins the behavioral level. Several other authors have proposed evolutionary models of holarchy (Ouspensky 1961; Land 1973; Jantsch 1980). The most comprehensive model of this kind I'm aware of is Wilber's (1995, 1997), though my view differs from his in several substantial ways (Smith 2001b, 2002b).
One of the most important implications of holarchy is that evolution creates complexity through new forms of social organization. Though as mentioned earlier, complexity is difficult to define, it's usually considered to be closely related to the number of different states that a system can realize or exist in. If each component of a system can exist in x different states, then the total number of states a system of n components may theoretically exist in is x raised to the nth power: xn. In other words, as more components are added, the complexity of the system increases disproportionately, a simple example of Changizi's scaling law.[26] Based on this logic, he and others have emphasized the close relationship of complexity with hierarchical organization (Simon 1962; Changizi 2001c; McShea and Changizi 2003; Valentine 2003).
When individual holons combine into social holons, however, it's not only the complexity of the entire assembly that increases. The complexity of the individual member holons may also increase. Put in simplified terms, this occurs because the more interactions an individual holon can make with other individual holons, the greater the number of states that it can potentially realize.
For example, in the simplest system conceivable, consisting of just two components, each component may exist in just two states: interacting with the other component, or not interacting with it (see Fig. 2). In a system of three components, each component potentially may exist in four different states, depending on which of the other two components it's connected to. In a system of four components, each component can exist in potentially eight different states, and in a five component system, each component may realize sixteen different states. Thus as new components are added to the system, the complexity of each individual component may double, representing an exponential increase.
Figure 2 represents a highly simplified scheme. In reality, the number of realizable states of individual components within a system may depend on many factors besides the number of such components.[27] Passing over such details, however, this relationship can be expressed as a very general rule: individual holons increase in complexity by becoming associated with increasingly more complex societies of such holons.[28]
To illustrate this point with an actual example, let's return to consider the visual system. I earlier described how individual neurons at various hierarchical levels of the visual pathway elaborate higher degrees of dimensions. An individual cell at one level of the visual pathway, where it integrates information from multiple cells at a preceding level, is more complex than these lower level cells. It experiences, or responds to, a greater number of dimensions, or a greater amount of detail in the external world. The increased complexity of the cell at the higher level depends critically on its connections with these other cells. For example, a cell in the V1 region of the visual cortex that responds to edges is connected to several or more cells in the lateral geniculate, which respond only to light intensity (Chapman et al. 1991). Its response may also be modified by inputs from other cells in the V1 region as well as outside of this region (Angelucci and Bresloff 2006). It is the input from all these cells that allows it to respond toand if we adopt the panpsychist view, experienceedges.[29] Thus the experience of two dimensions in this example is a social property, one that certain individual cells within a group or network of cells participate in.
Though individual holons in a society express complexity through their multiple interactions with other holons, generally no individual holon can exist in as many different states as the entire society can. In a two component system, the total number of possible states of the system is just two, the same as the possible number of states of each component (Fig. 2). But a three component system may exist in eight different states, depending on the connectivities among its individual components; a four component system may exist in 64 different states; a five component system may exist in 1024 different states. So while the complexity of individual holons may increase substantially as the society of which they are members increases in sizedoubling with each new component added in our theoretical example—the complexity of the entire society may increase by a far greater degree. Its rate of increase doubles, from two to four to eight to sixteen, going from a two component system to a five component system. In other words, the complexity of the entire system increases more than exponentially as a function of the complexity of individual components.
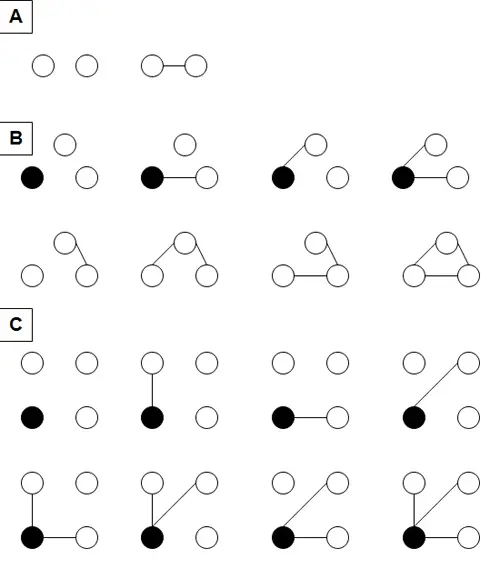
Fig. 2. Relationship between individual and social complexity. (A) In a system consisting of just two components, each component can exist in just two possible states, interacting with the other component, or not interacting with it. These are also the only two possible states of the system as a whole. (B) In a three-component system, each component can exist in four possible states. These are shown in the top row, for the component indicated by the filled circle. This three-component system as a whole, however, can exist in eight possible states, including the four shown in the top row, and an additional four shown in the bottom row. (C) In a four-component system, each component can exist in eight possible states. This is shown for the component indicated by the filled circle. The four-component system as a whole can exist in 64 possible states, which are not shown here. As discussed in the text and endnotes, this scheme has some simplifying assumptions.
Ignoring again real-life details, this relationship can be expressed as another general rule: while individual holons derive their properties from membership in some society, only society, taken as a whole, completely expresses these properties. In the example of the visual system, a single V1 neuron responds only to some feature of an object in the environment, such as an edge, or color, or movement; only a large population of neurons can put all these features together, resulting in the recognition of some specific object. This view is often expressed in the notion that there is no grandmother cellno single neuron in the brain that selectively responds to all of the features of one particular object or person. This reflects a widespread belief among neuroscientists that such complete responses are the properties only of large networks or populations of neurons.[30]
In this book, I argue that we can and should generalize this notionapply it not simply to neural activity in the brain, but to hierarchical processing at all levels in nature. Let's consider the physical level, which begins with atoms (Table 1). Though I have described atoms as zero-dimensional, this is actually true only for atoms that exist autonomously, outside of molecules. Examples of autonomous atoms include helium, as well as atoms like hydrogen and oxygen that can exist in very small molecules that function essentially like atoms.[31] Atoms like this may also exist within more complex molecules, however, and when they do, they may have a higher degree of dimensionality, which they acquire through their interactions with other atoms in the molecule.
For example, as will be discussed later at some length, a peptide molecule is a two-dimensional holon or structure. It results from the holarchical combination of amino acids (one-dimensional holons), which in turn are composed of individual atoms (zero-dimensional holons). A peptide molecule has many two dimensional properties, such as the ability to discriminate other types of peptides, and more generally, to recognize surfaces of other molecules. Some atoms in a peptide molecule, by virtue of their membership in this structure, participate in these two-dimensional properties. For example, they can distinguish between different classes of what is conventionally considered the same kind of atom. Thus a hydrogen atom in a peptide may interact with a particular oxygen atom that it is not covalently bonded to, and not some other oxygen atom. This is a two dimensional property. It is not exhibited by autonomous atoms, nor by atoms in small molecules such as individual amino acids.
Or consider the behavioral level, composed of organisms and their societies. Some insects exist in colonies or societies that are composed of several classes or castes, such as drones, queen attendants, foragers and food storers. These societies have a two-dimensional structure resulting from the holarchical combination of the several different classes or castes (one dimensional holons), which in turn are composed of individual organisms (zero-dimensional holons). These societies manifest many two-dimensional properties, such as the ability to 1) create multiple subroutines within vital tasks, such as feeding, reproduction and defense; 2) discriminate different types of others, including predators, prey, and allies, as well as members of the same species that are non-members of the society; and 3) create hives, mounds, and other home structures that are distinct from the rest of the environment.
By virtue of being a member of this social structure, each individual organism participates in these two dimensional properties. Thus individual insects can distinguish kin, or other members, from non-kin, as well as, within kin, members of different castes. Individual insects can also distinguish two physical dimensions of space, in other words, recognize many basic shapes and surfaces. These two-dimensional properties reflect a partial experience of the much more complete properties of the entire society.
The best known example of how individual complexity increases with social complexity, however, is our own species. All those features which most distinguish us from other animals, such as language, self-awareness, tool use, our ability to count, our understanding of concepts such as agency and causality, and our perception of time, are closely associated with our existence in more hierarchically developed societies. They depend upon complex interrelationships among individuals that could not exist apart from such societies.
Many animal behaviorists, in fact, now believe that the evolution of brains of other, non-human primates was critically dependent on social organization; that social interactions presented complex problems that could only be solved by evolving certain complex features of the brain (Humphrey 1976). Natural selection, argue the primate biologists Dorothy Cheney and Robert Seyfarth, has favored individuals who develop theories of social life.[32] My fundamental claim here is that this social intelligence hypothesis, as it has been called, is just the specific manifestation, among primates and other higher vertebrates, of a much more general rule that applies throughout nature.
The notion that evolutionary change takes place primarily on the social level may seem at odds with the conventional scientific worldview, but it's quite consistent with neo-Darwinism. Natural selection, as a widely used textbook emphasizes, involves changes in populations of organisms, which is to say societies of interbreeding individuals:
It is the population, not its individuals, that evolves, as some heritable variation becomes more common at the expense of other.[33]
While selection is commonly said to act on individuals, only societies have the complete set of individual properties or phenotypes on which selection can act; only societies incorporate all forms of evolutionary change. Moreover, the individual variations of organisms that are most likely to be selected are those that increase the number and nature of interactions they make with each other. I believe this is a critical point that neo-Darwinism has generally not been sufficiently aware of.
It's just because individual or fundamental holons have this ability to participate in the higher-order dimensionality of the stages they exist within that we can speak of different dimensionalities of these fundamental holons. That is, we can identify and characterize zero-dimensional, one-dimensional, two-dimensional and so forth properties of atoms, cells and organisms. So as we see in Table 2, we can rank not only social organizations, but also individual kinds of organisms, according to the dimensionality of their experience. (In a similar fashion, we can rank different kinds of atoms, and different kinds of cells). The stages of organisms shown in this Table, which we will examine in detail in subsequent chapters, correspond closely to the stages of social organization shown in Table 1C. To repeat, they have this dimensionality, according to my model, only because they participate in social organizations which actually manifest these higher properties.[34]
DIMENSIONS IN THE EVOLUTION OF ORGANISMS
| Dimensions | Representative stages | Examples |
|---|---|---|
| 0 | plants; primitive multicellular organisms | Volvox; Fungi; Cnidarians; Echinoderms |
| 1 | lower invertebrates | Nematodes; Annelids; Bivalve Molluscs |
| 2 | higher invertebrates | Crustaceans; Gastropod Molluscs; crawling insects |
| 3 | social invertebrates; lower vertebrates | flying insects; Cephalopod Molluscs; fish; amphibians; reptiles |
| 4 | higher vertebrates | birds; mammals |
| 5 | primates | great apes; humans |
Public and private experience.
So far, I have mostly described dimensions in structural and functional terms, resulting from the combinations of physical entities like atoms, cells or organisms into increasingly complex social organizations. But as I implied in the earlier discussion of the visual system, a key premise of my holarchical view is that higher orders of structural dimensions are closely associated with higher order experience dimensions. These general relationships are summarized in Table 3.
Note that for each hierarchical level, I distinguish two basic forms of consciousness, an exterior and an interior. This division corresponds fairly closely to what some theorists have referred to as environmental consciousness and self-consciousness, respectively (Mead 1967; Duval and Wicklund 1972; Carver et al 2002; Morin 2006). It may also be described in terms of perceptual consciousness vs. conceptual consciousness (Legerstee 1999), though as applied to non-human organisms, the notion of concepts does not strictly apply, and would have to be replaced by a somewhat more general term.
The exterior type of experience can be characterized most generally in terms of spatial dimensions, while the interior experience I describe as a self in relationship to the external world, and in particular in relation to other organisms. We can appreciate this distinction easily enough if we consider our own experience. We are conscious of an exterior world of three dimensions of space, populated with rocks, trees, cars, buildings, other people, and so on. Some philosophers also refer to this kind of experience as public, because it's one we all share. [35] We presumably all see much the same thing when look at a rock, a tree or a car, for example. [36]
THE DIMENSIONS OF EXPERIENCE
| Dimensions | Form of Perception | Functional Activity | Exterior (Public) Experience | Interior (Private) Experience |
|---|---|---|---|---|
| 0 | no awareness of space or time | autonomic, vegetative | no self/other distinction; no awareness of space or time | |
| 1 | aware of dualisms (e.g., front/back, up/down, left/right) | Reflexive, stimulus-response | distinguishes between self and other; intensity discriminations | I/thou relationship; primitive sign recognition |
| 2 | aware of surfaces | Kinesthetic, postural | distinguishes between different classes of others (e.g., predator/prey; kin/non-kin; male/female) | aware of itself as member of a group; primitive third person view |
| 3 | aware of 3-dimensional space | sensation, movement | distinguishes between individuals of its own species | aware of itself as unique individual |
| 4 | aware of 3-D space and linear time | feeling, emotion; behavior patterns | aware of behavioral states of other organisms | aware of behavioral states of itself |
| 5 | aware of time as extended and flowing | thinking, cognition, learning, memory | aware of permanence of objects and other organisms in space and time | aware of its own permanence in space and time; symbol recognition and manipulation |
But we are also conscious of an interior world of thoughts, ideas, feelings, imaginations, fantasies, and so on. This type of experience is closely related to our identity, because it shapes our sense of who we are—how we are similar to, and how we are different from, other people and other things.[37] This kind of experience is also called private, because it's different from one individual to the next, and can't be shared as easily or as completely as experience of exteriors can. This world also has a dimensionality, though here the dimensions are in the looser sense, as layers or stages. For example, as noted in the previous section, I consider the awareness of membership—that one belongs to a particular group—a second dimensional experience, and contrast it with the one-dimensional experience of distinguishing self from other. Just as two mathematical dimensions (a plane) includes one dimension (a line) while making a further differentiation of the kind of line (horizontal or vertical), the ability to distinguish kin from non-kin includes the ability to distinguish self from other, but makes an additional differentiation within the realm of other.
The distinction between exterior or public experience and interior or private experience is an old one in philosophy, going back at least to Descartes. The distinction is of course intended to apply only to human consciousness, but it's my claim, as shown in Table 3, that it can be extended to other forms of life. Though most other organisms presumably experience the world very differently from the way we do, I believe that the fundamental public/private distinction is preserved at all levels of existence (except in the lowest stage of any level, where the lifeform makes no distinction between itself and its surroundings).
This is a fairly strong and unconventional claim, and requires further elaboration. In his Critique of Pure Reason, Immanuel Kant argued that our exterior experience of the world is directly related to an awareness of (three dimensions of) space, while our interior experience presupposes an awareness of time:
Space then is a necessary representation a priori, which serves for the foundation of all external intuitions...Space is nothing else than the form of all phenomena of the external sense, that is, the subjective condition of the sensibility, under which alone external intuition is possible.
Time is nothing else than the form of the internal sense, that is, of the intuitions of self and our internal state. For time cannot be any determination of outward phenomena. It has to do neither with shape nor position; on the contrary, it determines the relation of representation in our internal state. [38]
Advances in science as well as philosophy since Kant may have cast some doubt on whether space and time can be separated so easily.[39] But without getting into these issues, I think we can conclude that an awareness of space is an essential feature of our public experience, and an awareness of time is critical to our private experience. As we will see, almost all organisms behave in a way that implies processing of both spatial and temporal information. Studies that we will examine in the following chapters demonstrate that they can recognize and respond to various features in the space around them; and that they can also recognize and respond to changes in these features, by learning new responses to stimuli, for example. I will actually argue in this book that most other organisms do not experience a flow of time as we do. Nevertheless, to the extent that this dimension plays a critical role in their experience, that experience is private.
The notion that most organisms have some degree of self-consciousness or self-awareness is certain to be controversial. While most scientists and philosophers accept that many other vertebrate organisms besides ourselves are conscious, self-consciousness is generally thought to be restricted to our own species, and possibly to a few non-human primates. However, it's important to understand that when I claim that even very primitive organisms may have some experience of self, I mean it in quite a different way from our usual understanding of self-awareness. When scientists, psychologists and other interested researchers discuss the self, they view it as identification with a unique form of life. Each of us is an individual, different from every other individual on earth, and it is from our experiences of this uniqueness that we develop our sense of self.
But it is possible in principle to have a much simpler sense of self (see Table 3). For example, most organisms, even very simple invertebrates, behave in a manner that suggests they distinguish themselves from the environment. To the extent that they have any awareness, therefore, they should have the experience of themselves as something apart from the rest of the world. This does not imply any sense of being unique, however, for that requires multiple distinctions drawn between themselves and other organisms. Likewise, the panpsychist view implies that certain more evolved invertebrates experience themselves as members of a group. This is a higher, more evolved sense of self, but it still lacks the experience of individuality, since the organism behaves more or less exactly like other members of its group. So as we will see throughout this book, as dimensions of experience of the outer world evolve, so do dimensions of the self that interacts with this environment. What we humans call our self-consciousness or self-awareness is, in this view, only the latest and most highly evolved form of self that organisms can be aware of.
SEE MORE ESSAYS WRITTEN BY ANDY SMITH
BOX 2
HIERARCHY VS. HOLARCHY
The word hierarchy, for many, evokes a rigid system of roles, such as traditionally exists in the military, the Catholic Church, and many government institutions. The nested hierarchies or holarchies found in nature, however, are somewhat different, and frequently natural hierarchies are, too. Hierarchies as they exist in human institutions consist of individuals who are at progressively higher stages within some social structure. Holarchies consist of the social structures themselves.
Hierarchy: It's lonely at the top. Hierarchy is frequently shaped like a pyramid, with the number of individual members progressively decreasing at higher stages.
Holarchy: The higher, the more inclusive. In holarchy, higher stages contain lower stages, so are necessarily larger.
Hierarchy: Control from the top down. In institutional hierarchies, individuals at higher stages may command or control the behavior of individuals at lower levels.
Holarchy: Bidirectional interactions. In natural holarchies, lower holons and higher holons influence each other. In protein molecules, for example, the overall conformation tightly constrains the positions of individual amino acids. But changes or modifications of individual amino acids can alter conformation. In the organism, the rates of growth and reproduction of individual cells are strictly regulated, but some individual cells control changes in the overall metabolic state of the organism. In human societies, certain forms of behavior are forbidden, but behavior by some individuals may result in changes in social organization.
Hierarchy: a linear chain of command. Hierarchical relationships can generally be traced from higher individuals to lower individuals in a sequential order.
Holarchy: Networks. Holarchies can exhibit complex relationships. For example, scale-free networks, which characterize many forms of human interaction (Barabasi 2002; Buchanan 2002), can form in a manner in which smaller networks are holarchically combined into larger networks (Ravasz et al. 2002; Ravasz and Barabasi 2003). Many randomly organized networks also form holarchies, for example, amino acid interactions within protein molecules. In fact, in nature, all networks are formed by the interactions of holons of some kind, and most networks are in turn holarchically combined into higher forms of life.
Hierarchy: Fixed Roles. Individuals in institutional hierarchies are defined by particular functions they fulfill in the organization.
Holarchy: Multiple states. Many natural holarchies feature different kinds of interactions among members. For example, metabolic networks within cells are holarchical organizations of enzymes, receptors and other biological macromolecules, each of which has a characteristic function. But such networks can adopt different states in which the functions of some members become more prominent, while those of others are reduced or eliminated. Within organisms, the roles of certain cells, tissues and other multicellular holons vary according to a variety of factors, both internal and external to the organism; in the brain, cells can take on new functions in response to injury of other cells. In all higher vertebrate societies, members adopt different roles in different situations.
Hierarchy: Power rankings. Individuals within a hierarchy can be distinguished according to how much influence they have. Those at one stage are ranked higher than those at a lower stage.
Holarchy: Egalitarian. In most natural holarchies, most individual members are equal or nearly equal in their functions and properties. In some holarchies, a few members are better connected to other members, and these better connections may result in enhanced properties. For example, the ability of individual atoms within a protein molecule to discriminate other individual atoms is restricted. In the brain, some neurons have more connections to other neurons than other members, and play a greater role in information processing. In human societies, there are frequently great disparities in wealth and power. But even in these examples, the great majority of individual members are roughly equal in their properties, functions or access to resources. Moreover, such inequalities that do exist among individual members of these holarchies are not necessarily a result of hierarchical organization per se. For example, they are featured in all scale-free networks, even those that are not hierarchically organized.
The Postmodern Organism
To summarize the discussion so far, I'm claiming that for every form of existence there is a close relationship between structural organization and experience, and more specifically, between degree of hierarchical development and degree of dimensions experienced. Here I want to discuss this relationship in a little more detail. In particular, I propose to extend two fairly well recognized features of human experience to that of virtually all other organisms.
Experience and interpretation.
Hierarchical processing of sensory information has the effect of gradually removing experience from the immediate environment. The organism does not experience the world directly, but as mediated through this processing. This progressive distancing from the external world, says Derek Bickerton,
is simply the price that is paid for knowing anything about the world at all. The deeper and broader consciousness of the world becomes, the more complex the layers of processing necessary to obtain that consciousness.[40]
These processing steps, however, do not simply clarify the worldallow the organism to experience it in greater detailbut also interpret it. All incoming sensory information is subjected to pre-existing biases or perspectives that limit not only what the organism can experience, but how it can experience it.
Bickerton, a linguist, was speaking largely of our own species, and as applied to us, this view should not be controversial. We all recognize that we can form concepts or mental representations that may have the most tenuous relationship with actual experience. Thus we can not only recall the past, but anticipate the future. We can not only imagine events that might happen, but events that could never happen. Through the process of representing the world with various symbols, then modifying and manipulating these symbols in various ways, we have a virtually limitless capacity to create within our minds worlds that don't actually exist.
But the really significant point here, perhaps less appreciated by the layperson, is that even when we believe we are interacting directly with the environmentsuch as when we are gazing at some natural object like a tree—we are actually doing so through concepts. Arguably, we never experience the world directlyin the sense of raw sensory data—but only at some distance removed, usually through our mental representations or interpretations of it.[41]
In this book, I will extend this notion to virtually all other organisms. I am not arguing that other organisms can form concepts or symbolsthough as we will see later, we humans are probably not completely unique in this respect. But my claim is that most other organisms, even fairly simple invertebrates, nonetheless interpret the world. Their interaction with the environment, like ours, is not direct, but mediated, with the result that they, too, experience something that is not actually there. Something that is created by their rudimentary minds.
Consider, for example, the lowly planarian, a member of the phylum Platyhelminthes, or flatworms. In some well known experiments conducted half a century ago, planaria were shown capable of forming a conditioned response between light and shock. When a light stimulus was repeatedly presented shortly before an electrical shock, the planarian learned to contract its bodysomething it normally does only in response to shockin the presence of the light alone (Thompson and McConnell 1955).
Clearly, the planarian's experience of light changes under these conditions. It no longer sees it in exactly the same way that it saw it before the light became associated with the shock. Yet the light stimulus itself has not changed. It is physically the same event it was prior to the association. In this very important sense, the planarian's experienceif we are willing to grant that it has any at allhas taken a significant step away from the immediate sensory world. It is not simply sensing the light, but interpreting it.
McConnell's studies were carried out in the artificial conditions of the laboratory. It is not clear how relevant they are to the organism's natural existence. But there are numerous other examples of interpretive perceptions by invertebrates that play a critical role in their lives. Many arthropods, particularly social insects, can discriminate among a large number of distinct but highly similar chemical substances, or pheromones (Howard 1993; Breed 1998; Lahav et al. 1999; Frohlich et al. 2001; Dani et al. 2004). They use this ability to determine the degree of their genetic relatedness to other individuals of their colony or society, to recognize receptive mates, and to identify and remember individuals with which they have engaged in aggressive acts. To do so, these organisms frequently must recognize complex mixtures containing dozens or hundreds of different compounds.
As I noted earlier, this is an example of what I call two dimensional perception or experience, the ability to recognize groups or classes of stimuli. Class distinctions like this are constructed, by picking out, or attending to, certain features of the environment while ignoring other features that are equally accessible to the organism's senses. Thus the organism is sensitive to certain chemical substances, but not others. These features are then grouped together, so that they function as a unitary stimulus or experience—such as male or female, which constitutes a higher-order class distinction. Many of these class distinctions, even in higher organisms including ourselves, result from processes that are hard-wired into the brain. Edward O. Wilson (1998) refers to such processes as epigenetic rules; they also correspond closely to what Terrence Deacon (1998) calls iconic forms of perception, and what biosemioticists simply call sign recognition. Organisms that are limited to this particular form of perception constitute the lowest level of Daniel Dennett's evolutionary learning scheme, and he refers to them simply as Darwinian creatures (Dennett 1995).
Making such distinctions, among chemical substances or among other organisms, constitutes an interpretation or perspective of the environment. It involves experiencing the world in one way instead of in some other equally possible way. This classification process, of course, is generally not arbitrary. A scientist would not say that the male-female distinction is just a construct of the nervous system (though as I will discuss below, some philosophers might). We regard it as a fundamental aspect of life, part of the real world, with organisms simply being favored by natural selection to recognize it.
Yet studies in which arthropods are trained to distinguish mixtures of just two or several chemical substances suggest that these relatively simple invertebrates have the capacity to group together any combination of individual stimuli that they recognize, and to distinguish one combination from another as well as from its individual components (Lynn et al. 1994; Derby et al. 1996; Hosler and Smith 1996). This capacity, in which a hard-wired ability is modified by learning, may play an important role in forming class distinctions that are not so obviously out there. For example, they may be used by certain insects and crustaceans to form social dominance orders (Tibbetts 2002; Gherardi et al. 2005; Moore and Bergman 2005) and by honeybees to identify flowers containing the appropriate nectar (Hosler and Smith 2000).
Deacon calls such learned forms of perception indexical relationships, and notes that iconic and indexical relationships form a hierarchy of experience or perception, culminating in our ability to use symbols.[42] Dennett refers to the invertebrates in which these learning processes first emerge as Skinnerian creatures. As I discussed earlier in connection with planaria, any learned association involves perceiving a stimulus in a new way. The fact that arthropods can learn to discriminate classes that they are not hard-wired to recognize implies not just a new interpretation of some sensory stimulus, however, but a perception of classes as something beyond any specific example of such a class. In other words, they are experiencing something that is not in the external world.
Experience is intersubjective.
Though many scientists may balk at any comparison made between the perceptions of simple invertebrates, on the one hand, and human language, on the other, the essential point that even fairly simple organisms have hierarchical structures processing information beyond immediate sensory stimuli should not really be controversial. This is basically what any central nervous system does. A second and bolder claim I make in this book, however, is that these hierarchical processes can't be understood as located entirely within the nervous system of a single organism. As summarized in Table 3, and as I emphasized in the earlier discussion, my view is that consciousness is primarily a social property, and as such is associated first and foremost with social organizations of organisms, and only secondarily is experienced by individual members of these societies. In making this argument, I am not necessarily claiming that there is a unified group consciousness (an issue I will address in the final chapter of this book), but that consciousness is distributed or intersubjective. As it is almost always experienced, it is associated with the interactions of at least two and usually many organisms.
Again, this should be fairly obvious for our own species. For us, the intersubjective web of relationships is associated almost entirely with language. Language evolved through human social interactions, and it is difficult to see how it could have been otherwise; as Wittgenstein (1958) famously argued, it has no use or meaning for isolated individuals. To the extent that our experience is language bound, therefore (and for many philosophers, as we will see later, there is no aspect of human consciousness that does not involve language), it appears that this experience must be first and foremost that of a social network, and secondarily that of its members. The language that each of us individuals uses is a property of the entire society. Every time we speakindeed, as I will discuss later, during virtually all of our thinking existencewe are reaffirming our membership in some society.
Intersubjectivity, however, is one of those terms that may mean many different things, depending on how strongly one believes it affects consciousness (de Quincey 2002b). To make my understanding of it a little clearer, it's useful to compare and contrast it with two current views of consciousness. The prevailing view among most scientists today, I believe, is that consciousness is an individual phenomenon, totally determined by, or at the very least localized to, some specific portion of the brain or some other nervous structure. This is commonly expressed in the view that there are neural correlates of consciousness, or NCCs (Crick and Koch 2003). As defined by philosopher David Chalmers,
An NCC is a minimal neural system N such that there is a mapping from states of N to states of consciousness, where a given state of N is sufficient, under conditions C, for the corresponding state of consciousness.[43]
In other words, the assumption is that any state of consciousnesssome specific experience of the external world, for exampleis associated with activity in some defined set of neurons. Generally speaking, if these neurons are active in the appropriate manner, one has the conscious experience. Indeed, a strong implication of this view seems to be that any human experience whatsoever could in principle be generated by stimulating the appropriate part of the brain.
Does language have neural correlates in this sense? Is there a specific set of neurons that is activated when we have the experience of using or understanding a particular word, phrase, sentence or longer linguistic construction? There is no question, of course, that there are fairly specific areas in the brain dedicated to language, and some studies suggest that particular classes of words or concepts may have locations somewhat distinguishable from other types of words (Mitchell et al 2003; Shinkareva et al. 2008). But as Deacon emphasizes, the distributed nature of languagethe fact that the meaning or meanings of any one word emerge only through its relationships with many other wordsseems to render fruitless any attempts to localize it:
it does not make sense to think of the symbols as located anywhere within the brain, because they are relationships between tokens, not the tokens themselves; and even though specific neural connections may underlie these relationships, the symbolic function is not even constituted by a specific association but the virtual set of associations that are partially sampled in any one instance. Widely distributed neural systems must contribute in a coordinated fashion to create and interpret symbolic relationships.[44]
The key phrase in this passage is virtual set of associations. It is virtual because it does not exist in toto in the brain of any single individual. Language, to emphasize again, is a social property, meaning that only society as a whole has a complete (or the most nearly complete) understanding of a language. The meaning of any particular word belongs only to the society.[45] Any individual, of course, experiences a meaning when he uses language, and perhaps we can talk about a neural correlate of that experience; but to the extent that that meaning can be shared and communicated with others, it only points or refers to the social meaning. And it is this social meaning that allows language to function at all, to prevent it from being Wittgenstein's impossible private language.
Very few species other than humans use language, of course. However, some philosophers and scientists have argued for a distributed form of consciousness based on non-linguistic social interactions outside of the brain (Wilber 1998; de Quincey 1998, 2000; Hargens 2001b; Thompson and Varela 2001; Hurley and Noe 2003). The basic argument, as summarized by Noe and Thompson (2004), goes as follows:
If the content of perceptual experience depends crucially on the environment, as well as on skillful motor capacities and capacities for directed attention on the part of the perceiver as a situated agent in the environment, then it cannot be assumed without argument (as the NCC programme does) that there is any such thing as a minimal neural substrate sufficient to produce conscious experience. Rather, the substrates of consciousness in particular of visual perceptual consciousness seem to cut across the brainbodyworld divisions.[46]
In other words, to the extent that experiencing the world involves acting on it and modifying it (Sherman et al. 1997; Liebert and Starks 2004), the correlates of that experience include aspects of the external environment. The environment, and particularly other organisms, is intimately involved in the generation of the experience. As the above quote indicates, this view, which some researchers refer to as embodied consciousness[47] (Thompson and Varela 2001; Noe and Thompson 2004), is generally presented as an alternative to the concept of an NCC. Its adherents claim support for it in studies of humans and other higher vertebrates showing that the parts of the nervous system activated by/associated with a specific form of perceptual recognition are not necessarily fixed, but can be modified by such factors as early development, learning, and environmental context (Hurley and Noe 2003). These kinds of studies have not been carried out in any lower vertebrates, let alone in invertebrates. However, I will discuss studies later that indicate that when even fairly simple invertebrates interact, dynamic processes frequently occur that are consistent with an embodied or distributed form of consciousness.
The view of consciousness that I present in this book is consistent, I think, with the essential tenets of both these views. On the one hand, I believe the notion of an NCC is valuable, and it may be that every conscious experience has some neural correlate (in lifeforms that have a nervous system). However, I also contend that the preoccupation with identifying NCCs obscures the much more important point that consciousness, as it is almost always experienced, is extended or distributed. It is part of a web of interacting consciousnesses, each of which is influenced by others. This understanding goes beyond the embodied view, for as we shall see, it is true even for the experiences of organisms that are not in dynamic interactions with any other organisms at a particular point in time. Just as language is something all of us carry around with us all of the time, and use to interpret the world in virtually every encounter with it, so I will contend, the perceptual processes of simpler organisms, though they evolved to facilitate their social interactions, are used to interpret every encounter with their external environment. And one major feature or outcome of this interpretive process is the experience of a certain number of physical dimensions.
I can summarize this view most easily by describing consciousness as intersubjective in three different senses. In the weakest sense, whenever organisms interact with each other, the experience of each is dependent on, presupposes, the other. In a second and somewhat stronger sense, when organisms interact dynamically with each other, or with other aspects of their environment, their experience may depend on ongoing behavioral adjustments they make. This is the embodied view of consciousness, and may challenge the notion of a straightforward NCC. Finally, in the strongest sense, I claim that just as language affects all our experience of the world, the iconic and indexical forms of perception of simpler organisms affect all their experience of the world. With some significant exceptions, which we will examine in the very next chapter, I claim that all the behavior of organisms, and all their experience to the extent they have it, is shaped by such relational processes. Indeed, one way of understanding evolution, on every level of existence, is through the development of increasingly more complex intersubjective relationships; dimensionality as I define it is a measure of intersubjectivity.
One final point deserves re-emphasis. Earlier I said that even if one wishes to dismiss the idea that very simple organisms have any conscious experience, the model I will discuss in this book has value for understanding behavior. The example of male-female recognition/classification by simple invertebrates illustrates this. My claim, following the panpsychist view, is that the male experiences the female as a member of a specific class. But even if it in fact has no actual conscious experience of this recognition, it still behaves in this manner. It engages in sensory processing that allows recognition of the female as distinct from other members of the same species, along with motor activity appropriate to this recognition.
We will see that this is the case throughout this bookthat functional or behavioral properties can be described in terms of dimensions. Thus I encourage readers who are not comfortable with the notion of panpsychismwho do not believe that simple forms of existence are in any meaningful sense conscious or able to experience themselves or their environmentto substitute for consciousness or experience more objective or neutral terms such as recognition, interaction or communication. Sometimes in this book I may emphasize the latter terms, while sometimes I will unapologetically use terms like consciousness or experience. Readers can decide for themselves where they come down in this debate.
Are dimensions real?
I said earlier that the notion that we humans interpret our experience is not controversial. In a limited sense, this is true. However, taken to an extreme, the notion of interpretation is closely associated with the postmodern school of philosophy (Derrida 1978; Rorty 1991,1998), and is commonly expressed by saying that there is no given, or reality existing completely separate and independent of our experience of it (Sellars 1956). This apparent denial of an objective reality makes postmodernism anathema to many philosophers and to almost all scientists[48], and even most postmodernists would not attempt to extend the notion to other forms of life. Yet it seems to me that a large body of evidence, alluded to above and to be presented in detail this book, supports at least a limited form of the postmodern view. [49]
According to the scientific worldview, three dimensional space (or four dimensional spacetime) is an independent feature of the universe. But if many forms of life experience their world in fewer dimensions, how can we be so certain that our three dimensional experience is not also a perspective, or an interpretation? Given that the dimensions of experience unfold through hierarchical organization, why should we believe that our experience is ultimate or real? In fact, I will provide further evidence that our own experience of the world, though much richer than that of most other forms of life, is nevertheless subject to the same kind of limitations. There may be additional dimensions that we ourselves are not aware of.
As I mentioned in the Prologue, there is currently great scientific interest in the possibility of additional dimensions of space. But most scientists believe that if such additional spatial dimensions exist, they are very small, and curled up. One reason for this is that if the dimensions were large, let alone infinitely so, they could be inhabited by visibly large objects or phenomena that would interact with our world in strange ways. We would be rather like Abbott's Flatlanders, who found the appearance and disappearance of three dimensional objects passing through their plane of existence to be miraculous. A fourth spatial dimension, say physicist Michio Kaku,
seems to violate common sense. If smoke, for example, is allowed to fill up a room, we do not see the smoke disappear into another dimension. Nowhere in our universe do we see objects suddenly disappearing or drifting off into another universe. This means that any higher dimensions, if they exist at all, must be smaller than an atom. [50]
There are several possible counter-arguments to this claim,[51] but the one most relevant here is to point out that Abbott assumed his two-dimensional creatures were confined to two dimensions. Likewise, in the previous quote, Kaku imagines that we are confined to three dimensions of space (while smoke or smaller forms of physical matter are not). Many of the organisms I will discuss in this book, though experiencing two (or fewer) dimensions, nevertheless move about in and generally inhabit the same three dimensional space that we experience. Even if they had notions of causality and other reasoning powers like ourswhich we will see they almost certainly do notthey would not witness anything unexplainable. To experience the world in two dimensions, or one dimension, or even in zero dimensions in this manner does not mean that objects and events in three dimensional space are invisible, or can move into a dimension where they are invisible. It simply means that they are experienced differently from the way they are experienced by us.
If this seems difficult to imagine, here is a simple illustration. Suppose you are blind and deafas some people are, of courseand in addition, have no sense of touch. Your only source of information about the world beyond comes from your sense of smell. Using this sense, you can locate sources of odors anywhere in three-dimensional space, simply by following your nose to the highest concentration of the source. To do this, however, you do not have to experience three-dimensional space. All you have to experience is one-dimensional space, in which you discriminate the intensity or concentration of the chemical substance.[52] As we will see in Chapter 3, there are in fact many fairly simple invertebrates whose interactions with the world are very much like this. There are also single-celled organisms, such as bacteria, that exhibit this kind of one-dimensional behavior, and even certain kinds of atoms in small molecules.
This is simply the Kantian view of dimensions. As noted earlier, Kant believed that three dimensional space, far from being the property of some underlying reality or things in themselves (noumena), was in fact projected upon the world by our very act of experiencing it. Rather like those goggles that used to be popular for movie-goers, which allow us to see details in flat surfaces as existing in three-dimensional space, our brains somehow imbue everything with existence in three dimensions.
This of course raises the question of what the world is really like. In the extreme postmodern view, as I noted earlier, there is nothing that the world is really like; every view of the world is a perspective, or an interpretation. Another way of stating this, though, might be to claim that the world really is dimensionless. Different forms of life carve up or impose on this dimensionless world a different number of dimensions. Higher dimensional experience may be more usefulin evolutionary terms, more adaptiveand might also be considered more real. I believe a very strong case can be made for the notion that evolutionarily higher organisms experience the world more realistically that lower organisms. But as we will see in the following chapter, life begins as dimensionless experience, and it may end that way as well.
As Above, So Below
In concluding this chapter, I want to point out that one of the most important implications of the holarchical view that I am presenting is that it strongly implies the existence of analogies between very different forms of existence. As I noted in the previous discussion, certain kinds of fundamentally similar properties are exhibited in common by molecules, by tissues, and by societies of organisms. A similar point has been made by some biosemioticists, who argue that fundamentally analogous processes involving recognition of signs and processing of information occur on multiple levels of existence (Hoffmeyer and Emmeche 1991; Brier 2006).
I don't want to overemphasize the degree of similarity. Molecules are obviously far simpler forms of life than tissues, which in turn are far simpler than societies of organisms. But to the extent that these analogies do exist, they reflect the fact that when holons, at any level of existence, combine into higher-order holons, they do so according to similar rules. They encounter, we could say, similar problems, and overcome these problems by devising similar solutions. So even though the basic units being combinedatoms, cells, organismsare profoundly different, the ways they interact may be highly similar. And at all levels of existence, researchers today increasingly appreciate that it is the interactions that must be understood (Tononi and Edelman 1998; Hartwell et al. 1999; Changizi 2001b; Montoya et al. 2006)
We will see many examples of this later in this book, but here I want to point out one of particular interest to our own species. Within the past decade, theorists have discovered that a great many human social interactions involve what are called small world networks (Barabasi and Albert 1999; Barabasi 2002; Buchanan 2002; Barabasi and Bonabeau 2003). These have been found, for example, in studies of the links between internet sites (Yook et al. 2002), email addresses (Ebel et al. 2002), literature citations (Bilke and Peterson 2001; Neumann 2001), language (Ferrer et al. 2001), acquaintances (Davidsen et al. 2002), sexual contacts (Schneeberger et al. 2004), movie actor roles (Ravasz and Barabasi 2003), trading partners (Andersson et al. 2003; Serrano and Boguna 2003) and airports (Li and Cai 2004), among a great many others. One of the signal features of such networks is that every member is connected, by relatively few steps or links, to every other member. This has been summarized by the catch phrase, six degrees of separation (Milgram 1967).
However, social interactions among humans are not the only kind that have this small world property. A very similar kind of organization has now been reported among cells in the brain (Hilgetag et al. 2000; Sporns and Zwi 2004; Achard et al. 2006; He et al. 2006; Bassett and Bullmore 2006) as well as among molecules in cells (Jeong et al. 2000; Wuchty 2001; Yook et al. 2004; Babu et al. 2004; Rao and Caflisch 2004). In my holarchical system, these three types of societieshuman, neural and metabolicform the highest stage on their particular level.[53] It is certainly no coincidence that three very different types of holons have formed similar types of social organization. In addition to shortening the communication times between distant members, small world networks, particularly a class of them known as scale-free, are highly resistant to random errors or attacks (Albert et al. 2000), potentially providing an immense evolutionary advantage.
Such analogies are not only suggestive of underlying rules that govern all social interactions at a certain stage of complexity. They raise some profound questions that I will address in more detail later. For example, if cells in the brain are organized in much the same way as humans in societies, does this mean the experience of such cells, to the extent that it exists, is in some respects like ours? If asked to name the form of life with experience most like our own, most people would probably name a non-human primate like a chimpanzee. Studies of such primates have indeed shown that they exhibit evidence of a relatively high intelligence, perhaps even including genuine if limited comprehension of language (Savage-Rumbaugh 1990). But while cells in the brain are presumably nowhere near as intelligent or aware as these higher vertebrates, the quality of their experience, to the extent it exists, might in some significant manner be more similar to ours. They are in fact the only form of life other than ourselves that interacts directly or indirectly with thousands of other holons like themselves on a routine basis. And while neurons have traditionally been regarded as little more than digital switches, older (Waxman 1972) as well as more recent experimental studies (Koch 1998; London and Hausser 2005), along with new theoretical considerations (Hameroff 1999; Edwards 2004; Sevush 2005) suggest that individual neurons can perform computational activities that might be regarded as the cellular equivalent of mind.
Another intriguing question raised by these analogies is: if our consciousness emerges from the way in which neurons interact in the brain, does a still higher consciousness emerge from the fundamentally similar way that humans interact in their societies? I noted earlier that according to a brand of philosophy known as functionalism, consciousness does not depend on the particular interacting units that are involved, but only the way they interact (Dennett and Hofstadter 1981; Dennett 1991; Hofstadter 2007). In the view of functionalists, therefore, neurons are not a necessary prerequisite for consciousness. Any kind of signal processor will do if there are enough of them and they are connected in the right spatial and temporal relationships.
While as noted earlier this extreme functionalist view is highly controversial and has its share of critics, no one questions that the organization of neurons, apart from their individual properties, plays a critical role in consciousness. In this light, I find it hard not to conclude that talk about an emerging global consciousness is not simply metaphorical. Human beings are, of course, not simply as good as individual neurons at processing signals, but far superior to them. We exhibit vastly more sophisticated and varied behavior in our interactions among ourselves. If more than six billion such holons are connected in much the same general organization as are the billions of far simpler neurons in a mammalian brain, how, one might well ask, could a higher form of consciousness not emerge? [54]
I will return to these issues later. For now, I just emphasize that since these dimensional stages do repeat themselves on every level of existence, the evolutionary story as I will tell it will not unfold strictly in the conventional sense, beginning with the first forms of existence, and proceeding, in a purely sequential and chronological order, to those following them. It's a story, rather, that must be told—and appreciated—multiple times, as it has occurred on multiple levels of the holarchy. It's a story of how atoms came to create cells; of how cells came to create organisms; and how organisms came to create life on earth as we know it now.
To make the presentation simpler and more coherent, however, I will in the following discussion heavily emphasize evolution on a single one of these levels—that beginning with the earliest multicellular holons or organisms, and culminating in our own species (and beyond). I have chosen this level because: 1) it is the one evolutionists are most concerned about; 2) the available evidence, from the fossil record and other sources, is much more extensive for this level than it is for lower levels; and 3) the degree or intensity of awareness of lifeforms on this level, to the extent they have any, is presumably much greater, and therefore more comprehensible to us, than is the awareness of lifeforms on lower levels. But I urge readers to keep in mind the cyclical nature of this evolutionary scheme, and will constantly remind them of it with discussions at the end of most chapters of the functional and behavioral properties of life on the cellular and molecular levels.
Endnotes
Prologue
1. Rudy Rucker's Spaceland (2003) and A.K. Dewdney's Planiverse (2000) are higher and lower dimensional stories, respectively, that are exceptions. The fourth dimension of space has also long intrigued artists. Picasso's cubism may have been a rather crude attempt to portray this dimension, in which all sides of a three-dimensional figure should be immediately accessible to the observer. The play Picasso at the Lapin Agile by Steve Martin explores the relationship of the artist to Einstein's work. See also Miller (2001) and Robbin (2006).
Chapter 1
1. Some philosophers who advocate this view prefer the term panexperientialism, to avoid the implications that lower forms of existence have mental properties that are anything like our own (see deQuincey 2002). Likewise, they prefer to argue that all forms of existence have experience, rather than consciousness. It seems to me difficult to argue that something can have experience but not consciousness; certainly it depends on how we define consciousness. In any case, I will use the term panpsychism here, and use consciousness and experience more or less interchangeably.
2. In a highly recommended article on the history of and the support for and against panpsychism, Seager (1999b) suggests that there is a third argument for it, which he calls the intrinsic nature argument. In its essence, this is the claim that whatever properties of any form of existence we observe must reflect some deeper nature, that is most likely to be mental or conscious in nature. This seems to me to be a difficult argument, and in my view not entirely separable from the genetic argument, but I leave it up to interested readers to decide for themselves.
3. Dualism has a much older philosophical history, but the version in prevalence today derives from Descartes. I'm equating mind and consciousness here, as this was done by Descartes himself. In his day, we must remember, we knew virtually nothing about the brain, and all mental phenomena seemed mysterious. Today, however, philosophers commonly distinguish between mind and consciousness, or define consciousness in several different senses. I will do this, too, later, but for now I follow the thinking of Descartes.
4. I'm glossing over some important distinctions for the sake of a quick summary of the traditional mind/body problem. Strictly speaking, dualism is contrasted with monism, the belief that mind and matter are both composed of a common substance. While materialism is the commonest form of monism today, there are other forms, such as idealism, of the kind made famous by philosopher George Berkeley (Darcy 1998). In Berkeley's view, everything is composed of, or is a manifestation of, mind, including what we call matter.
In addition, some philosophers hold a monistic view in which mind/matter differences are accounted for in terms of what is called property dualism. In this view, mind and body may be composed of the same substance, but represent very different properties of this substance, and so appear to be very different. If these properties are fundamental—that is, cannot be explained in further terms, just as mass, velocity and charge are fundamental—then mind and matter, though of the same substance, require no further explanation. This is discussed further below in the text.
5. Philosophers such as Thomas Nagel (1986) and Colin McGinn (1999) while agreeing with materialists that consciousness ultimately reflects material interactions, argue that our brain has inherent limits that make it impossible for us to ever discover and appreciate this basis.
6. But see the subsequent discussion on zombies.
7. I prefer the distinction functional/experiential to the more commonly used exterior/interior, because I do not regard consciousness as an interior property, or at any rate solely interior. Consciousness is not inside matter in any of the senses in which that term is normally used. In fact, as I will discuss later, consciousness has both exterior and interior aspects or manifestations.
8. Most of these authors, and particularly Wilber (1995), make it clear that they view evolution of human consciousness as part of a process that began with earlier forms of life that were also conscious. That is, the accept the notion that consciousness was present in some degree from the very beginning of life, indeed in what we commonly call inanimate matter. Wilber's comprehensive map of existence (1995,1997) makes this particularly explicit, by including lower forms of existence down to atoms, all of which are said to have an interior experience as well as exterior structure. However, Wilber does not describe the characteristics or evolution of these lower forms of consciousness in any detail, nor do, to the best of my knowledge, any of the other authors cited.
9. A start towards this approach has been made by what is generally called mixed method research, in which quantitative and qualitative data are combined. The mixed method concept has perhaps been taken furthest by Wilber (2006), whose Integral Methodological Pluralism or IMP proposes that knowledge can be obtained through eight different zones or perspectives. These include, for example, phenomenology or introspection, in which an individual directly examines the contents of her mind; structuralism, in which one individual examines the phenomenological reports made by others; hermeneutics, studies of social phenomena including approaches such as textual analysis, and various kinds of studies of group dynamics; ethnomethodology, involving various anthropological approaches; and several other approaches, including conventional scientific empirical studies. Many of these approaches can at least in theory be applied to non-human animals as well as animals.
While I believe the overall spirit behind IMP is valuable, I find several significant inconsistencies in the way Wilber has formulated some of his specific categories (Smith et al. 2006a,b; see also Smith 2003). These categories in fact derive directly from his AQAL (all quadrants, all levels) holarchical model, which I also have criticized at length (Smith et al. 2001a,b, 2002b). Still, some kind of approach along the lines of Wilber's is surely called for. For an application of IMP to current studies of animal experience and behavior, see Esbjorn-Hargens (2008). For a general discussion of the strength and weaknesses of approaches like Wilber's, see Murray (2006).
Some scientists and philosophers might argue that all purely phenomenological approaches are suspect, because of abundant evidence that people can be mistaken about their own mental processes or experiences. Dennett (1991) has proposed the term heterophenomenology to refer to empirical studies of phenomenological data. The rationale of this approach is that what individuals say about their mental processes can provide valuable insight into these processes, even if they are sometimes mistaken about what they claim to have experienced. The problem is that Dennett, and many scientists and philosophers, do not really regard phenomenology as a separate domain of knowledge, since they believe it can ultimately be reduced to empirically understood processes in the brain. Of course, nowhere in his work does Dennett discuss the possibility that mystical or spiritual experiences can offer insights into understanding consciousness.
10. Arthur Young (1976) also proposed a view of existence in which dimensionality played an important role. In some ways, his dimensions are comparable to those of Gebser, in that they correspond to different ways human beings have of knowing, or accessing information about, the world. However, one important way in which Young's model differs from both mine and Gebser's is that higher dimensions are not necessarily correlated with higher degrees of evolution or complexity. Thus while he identifies a movement of dimensionality from zero to three in going from light energy to organic molecules, the dimensions then return to zero during the evolution of molecules to humans. This implies that atoms and subatomic particles are in some sense more evolutionarily advanced than organisms, a conclusion no scientist I know of would accept. However, I believe there are many important features of Young's model, including the existence of stages within levels, which is also a critical aspect of my model, and the concept of symmetry. Though symmetry is less important in my model, I agree with him that types of symmetry are a key factor in distinguishing certain forms of life, and their experience. This point will be elaborated on in subsequent chapters.
11. The title of Gebser's book was meant to indicate that in the new emerging aperspectival consciousness, as he called it, there was an escape from linear or sequential time, a simultaneous awareness of past, present and future. In this sense, humanity was now able to access an additional dimension to the three of space. Again, this view is somewhat different from the conventional one of four dimensions, in which an awareness of linear time is the fourth dimension. I will have a great deal more to say about dimensions of time in this book.
12. The earliest form or stage of consciousness identified by Gebser, the archaic, might indeed be associated with a zero-dimensional view of the world in a fairly precise mathematical sense. As I will discuss later, a consciousness which makes no distinction between self and other can be described as zero-dimensional. However, I don't believe Gebser meant the term to be taken in this way, because he never calls attention to this kind of relationship; he does not point out how this kind of consciousness is like the zero dimension in mathematics. And in any case, even if he did intend it to be understood in this way, the earliest Hominids did not fail to distinguish themselves from their environment. This is a Romantic fallacy in my view. Studies of animal behavior (Hauser 2000) make it clear that even less evolved organisms draw important distinctions between themselves and their environment. This distinction is surely even greater for the genus Homo. I believe it's fair to say that our ancestors distinguished themselves from their natural environment far less than we do, but like virtually all living things, it was essential to their survival to make this distinction to some extent. I will discuss this issue later, in this chapter and in other parts of the book.
13. This is my interpretation of Gebser. Gidley (2007) claims that deep sleep, in Gebser's system, is associated with the magical stage and the dreaming with the mythic stage. If Gidley is right, however, this makes the problem raised here in the text even worse, with deep as well as dreaming sleep a characteristic of humans with brains identical to ours at a gross anatomical level.
14. Those familiar with Gebser's work may well find the notion that he viewed our ancestors as zombies as ludicrous. Yet his use of terms like deep sleep and dreaming, it seems to me, strongly implies just this conclusion. Any reasonable sense of these words that I can imagine implies an organism that is not at all aware of its environment, and which therefore must behave in this environment completely oblivious to this behavior and its consequences.
To be sure, some theorists, very notably Ken Wilber (2003), argue that both deep and dreaming sleep are not in fact lower states of consciousnessas all scientists would view thembut rather higher states, each identical to a certain state realized by mystics who have clearly transcended the mental or rational state. If we understood the sleep states in this way, our early ancestors would clearly have a rich inner life, and not be zombies.
However, I think this claim has been adequately rebutted (Smith 2001d; Edwards 2003b). I would just point out, for example, that many other animals sleep and may even dream (Louie and Wilson 2001), and we do not regard them as realizing a higher stage than our own mental-rational level when they do so. Furthermore, for Wilber to hold this view is clearly inconsistent with his general acceptance of Gebser's stages of consciousness, for this would constitute what Wilber himself first identified as the pre-trans fallacy (Wilber 1981), confusing lower, pre-mental or rational stages with higher stages that transcend the mental-rational level. Wilber very strongly emphasizesand I strongly agree with him here, and presumably Gebser would, toothat the states experienced by our early ancestors were very different from the state, whatever it is like, beyond the mental-rational. To be sure, the latter may involve integrating or incorporating in some way these lower states, but it is not at all simply a replay of them. This point will be discussed in some detail in Chapter 2.
Even accepting this pre-trans distinction, it should be noted that individuals in Gebser's dreaming state stage would not be true zombies, since they would have a rich inner life, a consciousness of something, whereas zombies in the philosophical sense do not. Perhaps Gebser also believes that deep sleep also involves some kind of awareness, though I have no idea what that would be. The essential point, though, is that when one is asleep or dreaming, one's consciousness has no consistent relationship with the world as apprehended in the waking, mental-rational stage. So the behavior of these individuals in this world, if they were to survive, would have to be automatic and unconscious in precisely the same way it would be for hypothetical pure zombies.
15. I want to address briefly another conceivable counter-argument to the problem raised by Gebser's view. In the view of many philosophers associated with what is called the postmodern school, there is no given, that is, no way that the world really is entirely independent of our perceptions of it (Sellars, 1952). Without going into the details of this argument here, I just note that some postmodernists might argue that our early ancestors did not in fact have to deal with the natural environment as we understand it. In this view, this environment did not yet in fact exist, because there were no modern humans yet who could perceive it in this manner. In this view, the deep or dreaming sleep they were immersed in was the only reality present.
However, the fact that we, today, can observe other animalsless evolved than even the earliest hominids—interacting highly efficiently with apparently the same environment we are aware of strongly argues against this view. Either these animals do see much the same given that we do, or they are zombies that interact with this environment without any conscious awareness of it. In other words, the environment that we are aware of today has been around for a very long time (which of course is the view of all evolutionary scientists), certainly long before the human species arrived.
I want to emphasize that in making this point, I'm not arguing that the postmodernist view is incorrect. What I'm saying is that the given (limited and ultimately unreal that it may be) that we recognize is essentially the same as the given that at least all other highly evolved vertebrates perceive. Major differences in the given must only be apparent between ourselves and much lower organismsand in fact, the purpose of this book is to document these differences.
16. An additional problem with Gebser's view is that the brains of our ancestors, at least as far back as 50-100,000 years ago, were essentially identical to ours in all their anatomical features. Once our species, Homo sapiens, emerged, no further evolutionary changes in the structure of our brains occurred, at a gross anatomical level. Given the highly plausible premise that there is a close relationship of consciousness to structure, it's very difficult to see how the same basic brain could be associated with very different degrees or levels of consciousnesses. Gebser's archaic stage is associated with hominids predating Homo sapiens, who had a distinctly smaller and probably structurally different brain from ours. However, his magic and mythic stages both emerged within the past 50,000 years, when Homo sapiens evolved, and thus both these forms of consciousness, together with the modern mental or rational, must be the product of essentially identical brainsthe same major structures containing the same kinds of neurons. Of course, there may well be micro-anatomical differences among these brains in the way neurons are interconnected, but such differences would also exist between any two individuals at the same stage, say, two contemporary human beings. The question is whether differences in consciousness as great as Gebser is proposing could really be associated with brains that are so similar structurally.
Similarly, Pinker (1997) argues that early humans had much the same mental-rational capacity that we today have, even if it was applied to somewhat different problems. Pinker, as an evolutionary psychologist, who looks for evolutionary explanations for human behavior, is undoubtedly impressed, as am I, by the notion that we would not have evolved the capacity for highly rational thought at this early time unless we in fact were using it. Pinker and other evolutionary psychologists would in fact probably reject the notion that there is a major difference in level of consciousness between ourselves and the earliest Homo sapiens. DiZerega (1996) has made a similar point.
17. Changizi (2001c), p. 3. I have simplified Changizi's view slightly. By scaling, he means that as we add more components to the system, we see a larger proportional increase in the number of expressions. We can't really test this notion with the systems I am describing, because however many atoms, small molecules, and so on that there are, the number is fixed. But the spirit of this definition is surely preserved in the notion that a few units at one level result in a larger number at a higher level.
18. I am beginning the discussion of hierarchy or holarchy somewhat arbitrarily with atoms. I will discuss subatomic phenomena, in particular the possible role of quantum effects in consciousness, in Chapter 11.
19. Most biological polymers are in fact linear chains of units like amino acids, sugars or nucleotides. Furthermore, some of these polymers may exist in planar forms (such as the b-sheet conformation of certain peptides), while still others take on an approximately spherical conformation (most enzymes). So one can to a significant degree describe molecules in cells in terms of lines, sheets and spheres. Nevertheless, the concept of definitions as I use it requires a broader, less specific meaning, particularly at other levels of existence.
20. Crick observed that it requires less energy for cells to synthesize large molecules modularly, that is, by stringing together similar smaller units, then to build them up atom by atom. The same principleenergetic favorabilitysurely is a major reason why holarchy is found at other levels of existence as well.
There are of course very small molecules (what I call simple molecules) such as water, carbon dioxide, and molecular hydrogen, oxygen and nitrogen. All of these are composed of just two or three atoms. However, I regard these as so simple as to be essentially like atoms in their properties. Though they have new or emergent properties that are the hallmark of a new stage or level of existence, these properties are much less different from those of the individual atoms than are the properties of amino acids, for example. See Smith (2000a) for further discussion of this point.
21. The exception to this rule are those select atoms in select amino acids that are directly involved in the catalysis. This is an example of a lower order holon existing in higher dimensions through its participation in the functions of a higher order holon. I will have much more to say about this below.
22. DNA and RNA molecules can reproduce themselves in the artificial conditions of the laboratory. But these conditions generally require numerous additional molecules, such as specific enzymes, that in nature are found only within cells. RNA molecules are now known that also have enzymatic activity, and they may have played an important role in prebiotic evolution (Cech 1987). However, cells are clearly required for large holarchical combinations of molecules to reproduce.
23. The mixed hierarchical structure of the cell is basically what allows it to bring different kinds of holons together, as is required for reproduction. Thus for DNA to reproduce itself, it must be able to associate specifically with certain enzymes. This would be virtually impossible if the DNA and the enzymes were organized in pure holarchical form.
24. I follow Wilber (1995, 1997) in making the distinction between individual and social holons, though there are many important differences in our classification of particular kinds of holons into these groups (Smith 2000b, 2001a, 2002a).
25. To be consistent with organization on lower levels, societies of organisms should be organized into a new fundamental holon. Presumably this new holon has not yet completely evolved or emerged. It's my assumption that it is, or will be, represented by the entire planet, understood not simply in a physical, biological or even mental sense, but as a complex form of life not completely understandable by ordinary human consciousness. This will be discussed in Chapter 10.
26. This is a theoretical maximum for the number of states, assuming that every possible state of any individual component can combine with every possible state of any other individual component. In reality, there are likely to be constraints. In fact, as made clear in the discussion below in the text, the maximum number of possible states of a system will usually be considerably less than this theoretical number. Still, as this same discussion shows, even with such constraints, a disproportionate increase in states occurs as the number of components increases.
27. In real systems beyond a certain size, individual components generally cannot interact with all other components, which will reduce their complexity. On the other hand, complexity may increase if the connections between components may vary in such parameters as strength and valence (i.e., positive or negative effect). Individual components may also access some states independently of connections with other components, through, for example, internal changes in their own component holons.
28. Anderson and McShea (2005) argue, on the basis of an extensive review of various behavioral and other capacities, that there is an inverse correlation of individual complexity and social complexity among various ant species. In general, members of small, simple colonies are generalists, able to perform several different forms of behavior, while members of large, complex societies are specialists, with highly restricted behavioral repertoires.
I believe, though, that this is a limited and somewhat misleading view of complexity. It's like arguing that our ancestors, who as individuals were directly responsible for satisfying all of their biological needs, were more complex than modern humans, who are highly specialized and dependent on social arrangements to satisfy most of these needs. Complexity in my view is better understood as the degree to which organisms can make discriminations among other individuals and other aspects of their environment. Anderson and McShea note that there is a greater degree of communication among individuals in more complex societies, probably reflecting a greater usage of cues and modulatory signals. To make these discriminations, the nervous system of the individual ant would have to be somewhat more complex, though to my knowledge this issue has not yet been addressed in a comparative study of different species of ants.
In any case, regardless of the relationship between individual and social complexity among ant species, the two clearly correlate when one takes a broader view. Human societies are more complex than those of non human primates, which are generally more complex than those of other vertebrate species, and vertebrate societies are generally more complex than those of invertebrates.
29. Neurons have conventionally been regarded as basically digital switches, capable of existing in only two states, on or off. However, recent studies, which I will discuss briefly later in this chapter, suggest a more complex view. Though neurons generally may respond to other neurons, via their synaptic connections with them, in only two wayseither sending out an impulse to other neurons or remaining silentthey can exist in a very large number of different internal states, depending on the pattern of activity coming to them from as many as tens of thousands of different synapses.
An analogy can be made with our own basic behavioral states, such as pleasure, fear or anger. Many different situations may trigger in us one of these basic emotions. Though the emotion is the same, the fact that we may experience it in response to different situations reflects a greater degree of complexity than would be the case if the emotion could be triggered by only a single cause (as is the case, or more nearly the case, for less evolved vertebrates). In the same way, the limitation of neuronal output to a simple on-off response should not obscure the fact that an enormous amount of information processing precedes this response.
In addition, some theorists now propose ways in which the complexity of inputs coming into neurons may be preserved in complexity of responses (Bieberich 2002; Sevush 2005).
30. It is now well established that single neurons in the brain can respond to specific categories of objects, including faces (Fried et al. 1997; Kreiman et al. 2000), and one study has reported cells that respond specifically to one particular face (Quiroga et al. 2005). However, these cells are not in the visual cortex, but in higher processing regions involving memory, suggesting that the cells are not responding to direct visual information per se. Moreover, technical limitations of the study preclude the conclusion that these single cells are completely specific for a single face or other object.
More importantly, even if single neurons may respond to very complex objects such as human faces, they are limited to a single such discrimination. Only a large network of such cells can recognize many, indeed, thousands, of different faces.
31. See endnote 21.
32. Cheney and Seyfarth (2007), p. 117.
33. Campbell and Reece (2008), p. 416.
34. This implies that experience is a primarily a social property alsothat societies have their own form of consciousness. To say that these properties are social is not to say that society behaves as a unified entity, however. It is only to say that none of the properties of any individual member can be understood except in the context of the properties of other members. There is no such thing as properties of isolated individuals, but only properties of interacting individuals. So there is not necessarily a single social consciousness, but the consciousness or experience of each member is intersubjective. This point will be discussed later in this chapter, and in Chapter 11 I will address the notion of group consciousness more substantially.
35. This distinction is not always clear cut. McFarlane (1996), for example, has argued that the outer/inner distinction is not exactly the same as the public/private distinction, because some kinds of knowledge or experience, such as mathematics, are inner yet public. Moreover, our own experience of both outer and inner worlds is so intimately dependent on concepts or mental representations, which as we will see result from social interactions, that in an important sense even our private experience is public. However, for the non-human forms of life that are the focus of most of this book, I believe that inner can be equated unproblematically with private and outer with public.
36. As I will discuss later, most organisms do not have direct, immediate experience of the world. Their experience of objects or other organisms is relational or contextual. It is shaped by prior experiences which are private as well as public. This is especially true of our own species, where our external experience is shaped to a large degree by ideas, concepts, prejudices, and so on. Nevertheless, we all share our public experience to a far larger degree than we share our interior, private experience.
37. Though the distinction between public and private experience is a convenient one, there are intermediate forms. For example, the kinesthetic or proprioceptive sense, which provides an organism with knowledge of both the position and the movement of its body relative to the external world, involves both types of experience. Sheets-Johnstone (1998) has argued that this sense is fundamental to the evolution of consciousness beginning with fairly primitive organisms. This is discussed further in Chapter 3.
38. Kant (1990), p. 23.
39. Indeed, a raft of thorny issues are raised by developments Kant never could have foreseen. The modern view that space and time form a continuum suggests that time, as well as space, is associated with the external intuitions. Haselhurst argues that quantum theory undercuts one of Kant's key assumptions about space, that it has no inherent movement associated with it. See http://www.spaceandmotion.com/metaphysics-immanuel-kant.htm
40. Quoted in Minsky (2006), p. 117.
41. Not all such processing involves language and symbolic representation, however. Perhaps the most primitive example of such higher order processing is found in color vision. The experience of color results from the brain's processing of wavelengths of light of different frequency. Physics tells us that the difference between wavelengths is continuous, along a spectrum. If we perceived color in immediate fashion, therefore, we should experience an infinite number of different colors. While we are indeed capable of discriminating a very large number of colors, we typically classify each color in some more fundamental group, such as red, blue or green. In other words, we have an innate tendency to carve up the continuous spectrum into a number of fairly discrete colors, albeit they are somewhat fuzzy at the edges. Each such group, such as red, blue and yellow, has a hue, called a focal color, which people consistently pick out as the best example of that particular color.
What might be called our digital experience of color results from the stimulation of three different types of receptorsfor red, blue and greenin the cone cells of the retina. The relative inputs from these three different receptors is transformed, in a higher processing stage in the visual cortex, into two axes or dimensions, one red to green, the other from blue to yellow. That is, every color is classified according to how much red (and correspondingly, how little green) it has, as well as how much blue (and how little yellow). (In addition, it is classified according to brightness). It is this coordinate system that allows us both to distinguish an enormous number of different colors while at the same time classifying all of them into a relatively small number of groups. The digital nature of this transformation has been particularly clearly illustrated in cross-cultural studies comparing the words for color in different languages (Berlin and Kay 1969). While different languages may recognize different numbers of colors, all are included in a few basic patterns. The simplest distinction is found between black and white (a brightness discrimination that I refer to as one-dimensional). Other languages add to this core, in sequence, red, green and yellow, blue and brown, and a few other colors. Thus there are languages with words for black, white and red, and other languages for black, white, red, green and yellow, but none that recognizes red without recognizing black and white, and none that recognizes yellow and green without recognizing black, white and red.
Further universal features of color vision have been demonstrated by studies in which people with a very simple color languagein which words for only black and white exist—were taught to name other colors (Rosch 1973). They found it much easier to name focal colorsbest examples of a particular part of the spectrum—than off center colors, taken from a different part of the spectrum. By showing that people of other cultures have much the same color experience as those with more sophisticated color languages, these results provide strong evidence against the notion that language determines color perception (and by inference, our recognition of other types of categories). That is, we have certain innate patterns which shape the kinds of words we use to describe our experiences, rather than the other way around. This point will be discussed further in Chapter 8.
Nevertheless, to the extent that we do tend to put colors into a few basic categories, our experience of colors is not immediate.
42. However, my view of these hierarchical relationships differs somewhat from Deacon's. The latter believes that iconic perception is the hierarchically lowest form of perception, followed by indexical and then symbolic processes. However, as suggested by the planarian, indexical processes may appear in organisms that lack iconic processes. This point will be discussed later, particularly in Chapters 3 and 4.
43. Chalmers (2000), p. 32. I believe Chalmers is making essentially the same point when he says elsewhere, It is widely believed that phenomenal properties depend only on the internal state of the subject (Chalmers 2004, p. 165).
44. Deacon (1998), p. 266.
45. As I will discuss in Chapter 9, in the postmodern view, every word has an essentially infinite number of meanings, since every time it is used it may appear in a slightly different context. With this understanding, we could say that only society possesses or accesses all the meanings of any one word (or at any rate, the maximum number of meanings). Notice also that this makes the concept of an NCC even more problematic. If a word means something slightly different every time it is used, there is presumably a slightly different neural correlate for it. But if this is the case, any particular neural correlate exists once and only once, which makes it virtually meaningless in understanding consciousness.
46. Noe and Thompson (2004), pp. 25-26.
47. The term embodied refers to the notion that consciousness is not localized to the brain, but involves other parts of the body. But to the extent that conciousness involves events outside the body, this term seems inappropriate to me. I prefer extended, distributed or intersubjective.
48. Many scientists felt their position vindicated when Alan Sokal (1996), a scientist, succeeded in getting published in a peer-review journal an article that was in fact a parody of the postmodern view. Desilet (1999), however, argues that developments in Sokal's own field of physics, beginning with Newton and continuing with relativity and quantum theory, provide striking parallels to the postmodern view of language, and offer evidence of their own that there is no escape from the interpretive web. Support for some version of postmodernism is also arguably provided by research in several other fields, including neuroscience (Damasio 1994; Globus 1995; Globus and Bezzubova 2001), cognitive science (Clark 1998), and linguistics (Lakoff 1987).
49. There is some irony to this, because the postmodernist school is often heavily critical of evolutionary theories such as the one proposed in this book. I will discuss this point in the final chapter.
50. Kaku (2008), p. 231.
51. In addition to the point made in the text, several others can be raised. First, we do of course experience many paradoxes or phenomena seemingly at odds with our notions of causality, such as quantum effects, in which observation or detection of a phenomenon changes its nature (Herbert 1985; Deutsch 1998), and the hard problem of consciousness itself (Chalmers 1996), which seems unexplainable by either materialism or dualism. Second, the ability to appreciate the very existence of a mystery may require experience of higher dimensions. Abbot assumed that his Flatlanders could understand causality, but as I will discuss in Chapter 7, an understanding of causality evolved only with ourselves and possibly some other higher vertebrates. It presupposes experience of three dimensions of space and one (or perhaps two) dimensions of time. In just the same way, the very limitation of our experience to a certain number of spatial and temporal dimensions may be associated with the lack of a critical feature of intelligence that would observe their effects in our dimensions.
Finally, there might be dimensions of time in addition to or instead of those of space that we are not aware of. I will discuss this in Chapter 7.
52. Actually, it would be quite possible to be sensitive to light, sound (vibrations), or touch also in this one-dimensional manner, and we will see some examples of organisms that experience the world like this. I use the example of smell because it is somewhat easier for us to imagine ourselves experiencing this sensation in one dimension than it is for sight or sound.
53. The analogous holon on the physical level should be the atom, not the large macromolecules which h have been observed to be organized into small world metabolic networks. In fact, both can be understood in this way. I will discuss these networks in more detail in Chapter 8.
54. In fact, the philosopher Ned Block (1978) used this idea as an argument against functionalism. He asked us to imagine a very large population, such as that of China, organized in the same way that neurons in the pain pathway of the brain are organized. Functionalism would predict that the Chinese nation would feel pain, a conclusion Block had no trouble rejecting.
The problem with Block's conclusion is that no individual human being can know whether in fact a higher form of existence feels anything or is conscious. Do individual neurons in the pain pathway feel pain? Presumably not; Block I am sure assumes that pain is only felt by the whole organism. In the same way, there is no reason to believe that any experience by a social organization of human beings would be accessible to individual members of that society.