|
TRANSLATE THIS ARTICLE
Integral World: Exploring Theories of Everything
An independent forum for a critical discussion of the integral philosophy of Ken Wilber
SEE MORE ESSAYS WRITTEN BY ANDY SMITH
DOES EVOLUTION
HAVE A DIRECTION?
Where Is It Going?
Andrew P. Smith
Wilber seeks to use the eros concept in places where evidence strongly suggests it's not needed.
David Lane is the most recent of several authors at this site to point out the flaws in Ken Wilber's view of evolution.* In doing so, he joins an even longer list of scientists, philosophers and other academics who—beginning with the celebration of the 150th anniversary of publication of The Origin of Species two years ago—have spoken out clearly and forcefully in support of modern evolutionary theory.
If there is one single aspect of this theory at the center of controversy, it is surely embodied in the word random. If even Einstein resisted the notion that the universe was a game of dice, it's difficult to blame lay people for trying to find design, or at least consistency, in the story of their origins. David Lane takes great pains to point out that randomness is only one aspect of Darwinism; natural selection, what Jacques Monod (1972) called necessity, seems to ensure that evolution has, if no purpose, at least some kind of direction. But if it has a direction, where is it going? If survival of the fittest is not a tautology, as some critics have claimed (Wilkins 1997), shouldn't evolution to some extent be predictable? Using evolutionary theory, we can look back into the past and understand how and why certain forms of life evolved. But what can we say about the future?
We need to address this question not simply to reassure people that life is more than just a matter of chance. Predictability goes to the heart of what science is and does.
Science can be succinctly defined as the attempt to identify the causes of phenomena. But the process does not stop there. When we think we know what causes a phenomenon, we attempt to use this knowledge to predict other phenomena, and ultimately, by causing them by our own actions. Indeed, scientific theories are generally validated only by the successful predictions they make. Einstein's relativity theory received an immense boost when it was demonstrated that the gravitational pull of celestial bodies did in fact result in the curvature of light. Conversely, modern string theory, for all its mathematical elegance, has failed to displace other so-called theories of everything (TOE) because of the difficulty of testing any predictions it might make (Smolin 2006)
Darwinism, nearly unique among widely accepted scientific theories today, has largely received a pass here. To be fair, the theory has proven, at the very least, to be quite consistent with many phenomena that were unknown when Darwin formulated it. For example, Darwin's ideas, when coupled with Gregor Mendel's experiments demonstrating the segregation and independent combination of hereditary properties, could have been said to predict the existence of genes. While heredity did not have to involve a macromolecule consisting of two helical strands of mutatable nucleotides, this structure turns out to have precisely the necessary properties. Moreover, comparison of DNA molecules of different but related organisms has shown that the further back in time these organisms began evolving separately—as estimated by fossil evidence—the greater their differences in nucleotide sequence (Cooper et al. 2003). This correlation between molecular and fossil dating, along with other fossil evidence of transitional or intermediate forms (Prothero 2007), constitutes powerful evidence for the theory, and should count as successful predictions. Indeed, as Theodosius Dobzhansky (1973) put it, nothing in biology makes sense except in the light of evolution.
Nevertheless, a key aspect of evolution is that it is not simply an historical process, but an ongoing one. It would of course be highly desirable that a theory of evolution should be capable of not just accounting for what forms of life appeared in the past, but for providing some insights into what might emerge in the future. This is where randomness becomes a problem. This randomness occurs not only in the gene mutations that are the source of hereditary variation, but perhaps also to some extent even in the selection process. The late Stephen Jay Gould, who referred to this latter kind of chance as contingency (1990), argued that random events in the past sometimes have had a major effect not just on what variants appeared, but on whether they survived. If we could rewind the evolutionary tape, Gould suggested—go back to any arbitrary date in the distant past and have evolution begin anew from that point—the results would be very different.
Like most evolutionary biologists, though, Gould was interested in the details of evolution—the specific types of structural adaptations developed by organisms. While it may be beyond our power to predict these in advance, what about more general trends? Perhaps there was nothing predictable about the emergence of a four-limbed creature that learned to walk upright and manipulate objects, but was the evolution of a brain, or some similar organ capable of processing information about the environment, inevitable? What about consciousness?
Here I will discuss the evidence for several major evolutionary trends, then briefly speculate on how they might extend into our future. As an important sidelight, I will make the case for the existence of several evolutionary processes other than Darwinism. The never-ending war between evolutionists and creationists has long tended to obscure the important point that one can fully believe in evolution without accepting that Darwinism accounts for all of it.
Complexity
Complexity is difficult to define, and several different definitions are in use. I will not attempt a precise definition here, but use an approximation that I believe is close enough to be useful. I define it in terms of the number of different states, or possibilities, that a system (a living thing or a machine) can exist in: the more states, the greater the complexity. This definition of complexity is, I believe, reasonably close to more precise definitions based, for example, on the number of computational steps required to create a system (Chaitin 1973; Bennett 1988; Lloyd 2007). In any case, it doesn't really matter if my definition fits exactly with these other, more precise definitions, because I am mainly interested in identifying a certain evolutionary trend, regardless of what name we give it.
We can easily appreciate my definition of complexity by considering what is often taken to be the most complex form of existence on earth: the human brain. With its billions of neurons and trillions of synaptic connections among these neurons, the brain clearly is capable of existing in an enormously large number of different states. Each state is distinguished from every other by the particular set of neurons that are active at that moment, and in the view of modern neuroscience, any particular such neuronal stateor at any rate, many of them—represents some specific form of information. Thus when we think or feel certain thoughts or emotions, a certain pattern of activity occurs in our brains, and this pattern is constantly changing as our thoughts and feelings change. Though the relationship of brain activity to thinking, feeling and other cognitive properties is not entirely understood, it's apparent that the enormous variety of behavior we are all capable of is closely related to the enormous number of different states our brains can exist in.[1]
Defined in this way, it seems obvious that there has been a major increase in complexity during evolution. If we consider the progression: small molecules, macromolecules, cells, invertebrates, vertebrates, humans—it's clear that over time forms of life have appeared that, by this definition, or indeed, anyone's reasonable definition, are more complex than any that preceded them. What makes this view controversial, though, is the implication that there is an inherent drive or purpose to evolution—shades of Wilber's eros—that produces this complexity. Some authors, like Robert Wright (2001), have embraced this idea, and find evidence for it throughout the history of both natural and human social evolution. Others, such as Gould (1997), have argued strenuously against it, conceding that while there has been some increase in complexity over evolutionary history, it results from chance, not any trend. In Gould's metaphor, evolution began with its back against a wall, representing zero or minimum complexity. Any change at all had to result in some increase, a step away from this wall.
I think the most reasonable position here—and the one probably accepted by the majority of scientists—lies between these two extremes. While there may not be an intelligent force guiding evolution to create life forms of increasing complexity, several scientific studies have provided evidence for an increase that does not appear to be a result of just the statistical fluctuations that Gould implied (McShea 1996; Adami 2002). David Lane, referencing Hermann Muller, points out that a process of gradually accumulating variations has the potential to create life forms of greater complexity. I would go further and say that an increase in complexity can often result in greater fitness, so that complexity is not simply possible, but often favored. Richard Dawkins (1997), one of today's strongest proponents of Darwinism, essentially made this point in a debate with Gould. Furthermore, other evolutionary processes that do not involve random variation and natural selection have also been described that could result in an evolutionary increase of complexity (Cavalli-Sforza and Feldman 1981; Kauffman 1993; Barabasi 2002; McShea 2005; Jablonka and Lamb 2005). I will be discussing some of these other processes later.
Those who argue against this idea, that natural selection very often results in increasing complexity, frequently cite the fact that some of the simplest life forms appear to be among the most well adapted. Bacteria, for example, have survived on earth for billions of years, and might well continue to survive under conditions that could result in the extinction of most or all multicellular organisms, including our own species. But those who make this argument generally overlook the fact that though bacteria are far simpler than any multicellular organism, they represent an enormous increase in complexity over what came before them—the primordial abiotic soup of organic molecules. In fact, most of the organic molecules found within bacteria and other kinds of cells today no longer exist on earth outside of these cells. They have been able to survive only by virtue of being part of a lifeform that can control, for example, the degradative processes that quickly destroy these molecules when they are found outside of living cells. So the complexity increase manifested in the evolution of bacteria, and in the still more complex eukaryotic cells that evolved later, represented, from the viewpoint of the cell's macromolecular components, a clear case of greater adaptive fitness.
What about a comparison of different multicellular organisms? There is a good argument to be made that, here, too, more complex life forms are frequently more adaptive. If our species goes extinct, it is most likely to be the result of a cataclysmic event that will also eliminate all other vertebrates and a great many invertebrates. Global warming, which along with other forms of pollution and habitat destruction is considered by many today to have the greatest potential to result in such a cataclysmic event, has already been shown to be a greater threat to many species considerably less complex than we are, including coral reefs and their associated marine invertebrate species and tropical rainforests, which are composed of an enormous diversity of plant and invertebrate as well as vertebrate species (Flannery 2005; Kolbert 2006; Wilson 2007). The World Conservation Union's Red List of endangered species as of 2007 listed 5742 vertebrate species, but also 2108 invertebrate species and 8447 species of plants. These numbers do not begin to tell the full story, as the status of the great majority of species, and particularly of invertebrate species, has not yet been evaluated; but it is telling that the number of threatened species as a percentage of species evaluated is far higher for both plants (70%) and invertebrates (51%) than for vertebrates (23%).[2]
To summarize, while there is no strict correlation between evolution and greater complexity, the fact remains that over time the process has created a significant number of different species which are more complex than any that preceded them. This has occurred not just once or twice or a few times, but literally dozens of times. While there are numerous examples of evolution going down paths that do not lead to greater complexity, it does seem that there is a bias in the selection process that ensures that greater complexity will be created more often than would be predicted on the basis of purely statistical fluctuations. From such a bias, can a trend be born?
Socialization
If natural selection seems capable of at least occasionally creating life forms of greater complexity, how exactly does it do this? It turns out that there is often a very simple answer: through combination of pre-existing life forms—whether they be molecules, cells or organisms—into societies. Such combination can potentially result in an enormous increase in the number of possibilities or states.
How societies become more complex
This can be demonstrated very easily mathematically.. Suppose we begin with a life form—what Wilber, and others including myself would call a holon—that can exist in just two different states. For example, a nerve cell might either be active, sending an electrical impulse down its axon, or inactive. If we combine this neuron with a second neuron, the combined system can exist in four different states—both neurons active, both inactive, or one or the other active. More generally, if a system consists of n components or holons, each of which can exist in x different states, then the total number of states that system may theoretically exist in is x raised to the nth power: xn. In other words, as more components are added, the complexity of the system increases exponentially.
However, the complexity increase in real-life social systems is potentially much greater than even this. Suppose that a holon adopts a different state every time it interacts with another holon. Then as new holons are added to the system, the number of states that both they, and the existing member holons, can exist in increases. This is shown in Fig. 1. For two holons, the system can exist in only two states, depending on whether or not the holons are interacting. For three holons it can exist in eight different states, while a system of four holons can exist potentially in sixty-four different states. The complexity or number of states increase does not follow the simple xn rule I mentioned previously, because the states of any one holon are not completely independent of the states of all other holons. But this may be more than compensated for by the increase in potential new states as holons are added to the system.
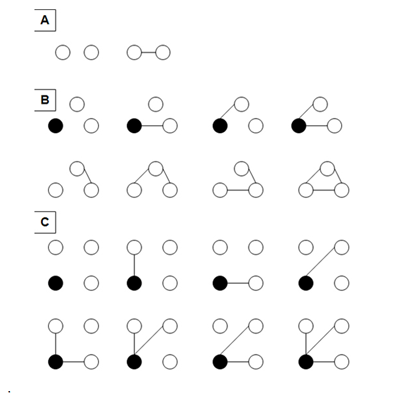 . .
Fig. 1. Relationship between individual and social complexity. (A) In a system consisting of just two components, each component can exist in just two possible states, interacting with the other component, or not interacting with it. These are also the only two possible states of the system as a whole. (B) In a three-component system, each component can exist in four possible states. These are shown in the top row, for the component indicated by the filled circle. This three-component system as a whole, however, can exist in eight possible states, including the four shown in the top row, and an additional four shown in the bottom row. (C) In a four-component system, each component can exist in eight possible states. This is shown for the component indicated by the filled circle. The four-component system as a whole can exist in 64 possible states, which are not shown here. As discussed in the text and endnotes, this scheme has some simplifying assumptions.
So the first lesson here is that complexity is created by social organization. Societies—and I use this term very generally, to mean not simply human or even animal societies, but combinations of molecules and cells as well—are generally more complex, and frequently, far more so, than any of their individual members. This is a point that has been lost not simply on Wilberwho clings to the erroneous and easily disproven idea that societies always have the same degree of complexity (and level of hierarchical development, an issue I will address shortly) as their individual members—but to some extent, I believe, on the entire evolution community.
The really important changes in evolution almost always involve combinations of holons into societies.
How individuals become more complex.
When individual holons combine into social holons, however, it's not only the complexity of the entire assembly that increases. The complexity of the individual member holons may also increase. Put in simplified terms, this occurs because the more interactions an individual holon can make with other individual holons, the greater the number of states that it can potentially realize.
This is also shown in Fig. 1. In the simplest system conceivable, consisting of just two components, each component may exist in just two states: interacting with the other component, or not interacting with it (see Fig. 1). In a system of three components, each component potentially may exist in four different states, depending on which of the other two components it's connected to. In a system of four components, each component can exist in potentially eight different states, and in a five component system, each component may realize sixteen different states. Thus as new components are added to the system, the complexity of each individual component may double, also representing an exponential increase (though the exponential power is always lower than that for the entire society).
Figure 1 represents a highly simplified scheme. In reality, the number of realizable states of individual components within a system may depend on many factors besides the number of such components.[3] Passing over such details, however, this relationship can be expressed as a very general rule:
individual holons increase in complexity by becoming associated with increasingly more complex societies of such holons.
Socialization and evolution of the brain.
I claimed a moment ago that all or most of the really important evolutionary changes result from formation of societies. Consider, for example, the evolution of new phyla, organisms which differ from their ancestors not in just a few structural details, but in a profound re-organization of the entire body. Critics of Darwinism have long argued that the theory has great difficulty explaining such major changes by a slow, gradual process. Without addressing the truth of this claim, I note that these kinds of changes are one of the primary results of socialization—which as I will discuss later may to some extent involve non-Darwinian processes.
The most important evolutionary changes underlying the emergence of new phyla occurred in the nervous system. In the transition from simple invertebrates to more advanced invertebrates to the first vertebrates to mammals, and finally to our own species, there has been a progressive increase in the size and complexity of the nervous system. This has not been the only evolutionary change, of course, but it's the critical one. How has this come about?
The short answer is through the association of nerve cells into ever larger and more complex societies that we call brains. Larger, more highly connected brains can process information that smaller, lesser connected brains cannot. Moreover, organisms with these more complex brains can engage in more interactions with other organismsin other words, form more complex societies and ecosystems. Indeed, it is fairly widely accepted that for not only humans, but other primates, brain and social systems co-evolved. A correlation between primate brain size and size or complexity of societies has been observed (Dunbar 2003), and according to the social intelligence hypothesis, a major driving force of primate brain evolution was the need to be able to process increasingly complex social relationships (Humphrey 1967; Reader and Laland 2002; Cheney and Seyfarth 2007; Holekamp 2007). As I will discuss in the following section, studies of other organisms, including invertebrates, suggest the existence of a similar relationship between the complexity of brain or nervous system and that of society. To the extent this occurs, we can conclude that the association of holons at two levels of existencecells into brains and organisms into societieshas been the primary driving force of the evolution of more complex organisms—not simply new species, but larger distinctions, up to and including new phyla.
Hierarchy/Dimensionality
I explained in the previous section that not only does complexity increase when several or more individuals associate into a society, but that the complexity of each individual can also increase. If an individual has multiple different interactions with other individuals, it can exist in multiple different states.
In order for this complexity increase to occur, however—at both the group and the individual level—the individual must recognize that each other individual is in some way different from every other individual. Only in this case can each recognition event be considered a different state. In the most primitive animal societies, represented, for example, by colonies formed by very simple invertebrates, this kind of recognition almost certainly does not occur. A large body of evidence indicates that individual organisms at this evolutionary stage do not distinguish among other individual organisms (Hauser 2000; Griffin 2001). Any one individual is essentially like any other.
This means that the level of complexity of such societies, not to mention that of their individual members, is very low. If there is no meaningful distinction between interaction of member A with member B and interaction of member A with member C, D, E, and so on, then all these different interactions effectively count as a single state, not different states.
Dimensionality: measuring the hierarchical development of societies.
This situation first began to change with more advanced invertebrates, such as arthropods. While relatively few of these organisms can distinguish among other individuals of their species[4], most of them can distinguish among different classes of individuals. This is particularly apparent among (though by no means restricted to[5]) the social insects, societies of which typically include several different castes, such as drone, forager, food storer, and queen attendant. Each member of the colony belongs to one, and only one, caste, and plays a specific role appropriate to that caste. And most important, in the context of the current discussion, each member of the colony can generally distinguish other members of its own caste from members of the colony that belong to a different caste. This is what I mean by class distinctions.[6]
A simplified comparison of these two kinds of societies is depicted in Fig. 2. The structure of very simple invertebrate colonies is homogeneous; each member is indistinguishable from every other member, and each member's interaction with another member is indistinguishable from any other individual-individual interaction. The structure of colonies of social arthropods, in contrast, is differentiated; it contains subgroups, which divide the membership into separate enclaves. Members of any subgroup interact differently among themselves from the way they interact with members of another subgroup.
I refer to simple invertebrate colonies as one-dimensional, because there is one hierarchical stage of development beyond the individual organism: the entire colony. I refer to arthropod colonies as two-dimensional, because there are two hierarchical stages. First, individual members associate into castes; second, the castes associate into the entire colony.
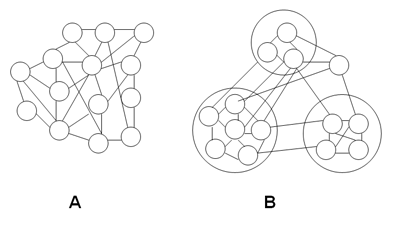
Fig. 2. One-dimensional and two-dimensional societies. In one-dimensional societies (A), the relationship of every member (represented by the small, inter-connected circles) to every other member is the same. In two-dimensional societies (B), members belong to different subgroups. The relationship of members of the same subgroup to each other is different from their relationship to members of other subgroups, and members must be able to detect these differences.
Thus dimensionality, as I define it, is a measure of hierarchical development, with each succeeding dimension corresponding to a new hierarchical stage. Though I have described only two dimensions, there are additional ones accessed by many organisms, which I will discuss briefly a little later. Because new hierarchical stages result in further differentiation among individual members of the society, they are associated with greater possibilities for individual interactions and increasing complexity. Hierarchy is of course a central feature of Wilber's entire system of thought (1995), but its evolutionary significance has also been recognized by others (Mayr 1982; Allen and Starr 1982; Raff 1996; Depew and Weber 1997). In particular, several scientists have emphasized the close relationship of complexity with hierarchical organization (Simon 1962; Changizi 2001c; McShea and Changizi 2003; Valentine 2003).
One- and two-dimensional perception by organisms.
This difference in the complexity of the society is reflected in the complexity of the individual members. For members of simple colonies, interaction is largely limited to determining the presence or absence of another member of the colony, without making any distinction between different members. That is, each member can only distinguish self vs. other, itself from some other member. I refer to this as one-dimensional perception or experience, because it can only make one kind of distinction. Members of arthropod societies, in contrast, possess two-dimensional experience. They can distinguish not only self vs. other—the first dimension—but also different kinds or classes of others. This is the second dimension.
Let's look a little more closely at how these dimensions are actually manifested in these organisms. A fundamental property of most simple invertebrates is the ability to make intensity discriminations. For example, these organisms can sense the strength of stimuli such as light, pressure, temperature or certain chemical substances (Rankin 2005; Bargmann 2006; Bounatas and Chalfie 2007). Since the perceived intensity of a stimulus is inversely related to the distance of its source from the organism, they can use this form of perception to locate and interact with other organisms. Such organisms actually sense or experience a single dimension of space, formed by a line or axis joining themselves to the source of the stimulus. The length of this line or axis changes as they approach or move away from the stimulus, and they judge this length or distance by the intensity of the stimulus.
This perceptual ability is the key to interacting with other members in a simple, one-dimensional colony, or even with organisms of other species. For example, the leech, a typical one-dimensional organism, locates its hosts by the decrease in light intensity caused when it approaches them (Bielecki 1999). Upon physical contact, further behavior is guided by temperature sensors that lead it to the host's blood supply, and perhaps also by chemoceptors sensitive to components present in the blood. These intensity discriminations, simple as they are, suffice to trigger most of its most important behavior.
Most arthropods, in contrast, sense two dimensions of space (though some, such as the flying insects, can sense three-dimensional space). This has been demonstrated in detailed studies of both ants and crabs, for example, which estimate the planar distance between two points that they traverse (Wohlgemuth et al. 2001; Walls and Layne 2009). Foraging desert ants, in particular, must estimate distances they have traveled fairly precisely, in order to return to their nest after they find food; but this distance is measured entirely in two dimensions, even when they must climb up and down intervening hills. Other evidence for two-dimensional perception comes from studies of the praying mantis, which will mistake for prey and attack simple cardboard rectangles, if they have the proper proportions of length and width (Prete 1990, 1992). As I have discussed in detail elsewhere (Smith 2009), experience of two dimensional space is ultimately derived from the ability of these arthropods to distinguish different classes of stimuli.
In summary, there is a significant relationship between the hierarchical development of a society and the number of spatial dimensions that its members can perceive or experience. Though I have discussed only two dimensions, there are several more. Some invertebrates, and most if not all vertebrates, are members of three-dimensional (or higher) societies, and can perceive or experience three-dimensional space. The higher vertebrates, birds and mammals, are members of four-dimensional societies, and experience three dimensional space as well as a fairly extended dimension of time. Our own species lives in five-dimensional societies, and we experience three dimensions of space, and have a unique experience of time that in an important sense is two-dimensional (Smith 2009).[7]
The embodied view of perception and consciousness
This is a very powerful claim that I'm making here, so let me make it quite clear. Societies result from the association of individual organisms, and these societies exist in different dimensions, determined by how many hierarchical stages they contain. A simple, homogeneous society is one-dimensional; a two dimensional society contains subgroups; higher dimensional societies manifest further differentiation in space, and eventually, time. Individual members of societies experience a corresponding number of dimensions as the society, and one (though not the only) manifestation of this experience is physical space and, in higher vertebrates, time. In other words, I'm claiming that the experience of space and time by all organisms, including ourselves, results from the complexity of the societies we are members of.
In the standard scientific view, of course, our experience of space and time, along with objects, organisms and events occurring within this space and time, is determined entirely by processes in the brain. While these processes reflect what is occurring in the environment around us, the activity of the brain, in and of itself, suffices to create the experience. The presumption is that every experience that we have is ultimately the result of some specific pattern of activity in the brain. Philosopher David Chalmers (2000) refers to this activity as a neural correlate of consciousness (NCC).
This view, however, has been recently challenged by several philosophers and scientists (Lakoff and Johnson 1999; Thompson and Varela 2001; Noe and Thompson 2004). These scholars argue for a so-called embodied view, in which perception, experience and consciousness have a more dynamic basis, emerging from ongoing interactions of the organism with its environment. While activity in the brain is still considered critical, it is envisioned as part of a larger pattern that includes the organism's entire body (hence the term embodiment) constantly adapting in response to changes in its surroundings. The whole organism plus its environment, one might almost say, constitutes the brain in which the correlates of consciousness are to be found. This view finds some experimental support in studies of both humans and other mammals (Bach-y-Rita 1972; Hurley and Noe 2003).[8]
The embodied view is likely to seem strange and counter-intuitive, at least to a scientist. Isn't all experience—not reality, mind you, but our experience of reality—ultimately localized in the brain? But there is one example of embodiment of experience that I believe nearly everyone accepts: language. While our ability to use language is mediated by specific areas of the brain, I think most of us would agree that the meanings of words are not localized to any individual's brain. They emerge from the social interactions of individuals, and in a very real sense exist only in the entire society or community of individuals. While there presumably are neural correlates of language, in the sense that specific parts of our brain become active when we use certain words or terms (Mitchell et al 2003; Shinkareva et al. 2008), the ultimate referents of these terms—what allows language to be shared at all—are found only in intersubjective processes.[9]
One way of understanding the embodied view—at least the version of it that I'm proposing—is to say that all perception or experience of the environment, by all organisms, is something like a shared language.[10] While experience of space and time certainly must have correlates in the nervous systems of these animals, the embodied view claims that this experience is ultimately social. It could not exist in the absence of a community of organisms. Just as language is created by individuals communicating with each other, experience of space and time is created by individuals behaving in various ways in relationship to each other (and in relationship to non-living things in the environment).
Are there additional dimensions that we can't experience?
This view has several profound implications, which I will be discussing as we go along. First, it suggests that our experience of the world as three dimensions of space and one (or more) of time is not necessarily the way the world really is (if is even meaningful to speak in this manner). Since many, in fact, the great majority of, species cannot experience, as we do, three dimensional space and at least one full dimension of time, one might reasonably ask whether we are in fact experiencing all the possible dimensions of our world—or whether further evolution could result in experience of still more dimensions.
I will return to this question later, but here I want to address a standard scientific objection. While string theorists claim there may be as many as ten or eleven dimensions of space, all of these dimensions beyond the familiar three are thought to be very tiny and curled up. Very few physicists believe there is a fourth, infinite dimension of space, or a second dimension of time.[11] A typical argument against this is presented by physicist and popular author Michio Kaku:
A fourth spatial dimension seems to violate common sense. If smoke, for example, is allowed to fill up a room, we do not see the smoke disappear into another dimension. Nowhere in our universe do we see objects suddenly disappearing or drifting off into another universe. This means that any higher dimensions, if they exist at all, must be smaller than an atom. [12]
Kaku, like most scientists, is assuming that we are confined to the three dimensions that we experience. That we are unable to enter or move about in any higher dimensions. He is echoing the same argument made by Edwin Abbott, in his classic, Flatland (1992), where two-dimensional creatures would have been mystified by the sudden appearance and disappearance of objects that existed in a third dimension that was inaccessible to the Flatlanders.
But there is no reason to suppose that, if there are additional infinite dimensions of space (or time), that would be the case. This is not the case for the simpler organisms whose experience or perceptual capabilities are limited to one or two (or even zero or a half) dimensions. We know that they exist not in some two-dimensional Flatland, or one-dimensional Lineland, but in the same three dimensions (at least) of space that we do. An enormous amount of behavioral data demonstrates that it is clearly possible to exist in more dimensions of space than one is actually able to sense or experience. The organism's perceptual apparatus simply collapses all the existing dimensions into two, or one, or however many it is capable of sensing.
If this possibility has not yet been taken seriously by most scientists, there is a good reason for it. There appears to be no way to test for it. If, for example, the universe really has four infinite spatial dimensions, but because of the limits of our sensory apparatus, we can only experience three of them, this would not change any of our current observations. Since everything in this four-dimensional world would be interpreted as a three-dimensional experience, the laws that we observed would be identical to those observed in a three dimensional universe. To be sure, if we were suddenly given the capacity to experience all four dimensions, the world would look very different—just as it would to a praying mantis that could see its prey in three dimensions, rather than as flat rectangles. But in an important sense, there would be nothing in this world that we had not seen before. There would still be rocks and trees and other animals and people, and all the products of our technology. We would just see all of these things much more fully.
Genetic selection and social selection
To summarize the discussion so far, we have seen that a major evolutionary trend involves individual holons—molecules, cells, organisms—associating into societies. These societies then differentiate into various kinds of subgroups, forming increasingly complex hierarchies. The greatest complexity is associated with the entire society, but individuals, by virtue of their membership in the society, also experience increasing complexity.
How or why does this socialization process, and formation of hierarchies, occur? Does it involve the Darwinian processes of random variation and natural selection? It may to a certain extent. Changes that occur at the cellular level, in the brains of organisms, for example, can result from mutations that change either the properties of individual neurons and/or their ability to communicate with other neurons. Changes that occur at the organism level, in animal societies, can also be selected for, because as I discussed earlier, the increase in complexity of societies is accompanied by an increase in the complexity of their individual members. In fact, membership in a group often offers immense fitness advantages, such as access to pooled resources, specialization of various tasks, and protection from predators. So there is usually no need to invoke group selection, a phenomenon that most evolutionists now believe accounts for a relatively small amount of evolution (Wilson and Sober 1998).
However, other, non-Darwinian processes may also be necessary for socialization. Consider the evolution of human language. We know that human beings have certain areas in their brain essential to the expression and understanding of language, and that all other species lack these areas, at least in as well developed form as found in Homo sapiens (Deacon 1998; Preuss 2001). Natural selection was almost certainly involved in the evolution of these brain areas. But we also know that language, as it exists today, is the product of more than just natural selection. As I noted earlier, the meanings of words cannot be located in any individual brain, but only arise from the interactions of many different individuals. Some, though not all, linguists believe that other, more basic aspects of language may also be learned (Pinker 2007). Such intersubjective processes cannot be the result of Darwinian evolution, because they involve the concerted behavior of large numbers of individuals. Darwinism acts to select the fitness of particular individuals, not the interactions they have with other individuals.[13]
A different kind of process, called cultural evolution (Cavalli-Sforza and Feldman 1981), was certainly involved, as well as other processes, one of which I will discuss later. Cultural evolution in fact has played a far greater role in our evolution than just the creation of language. Anything that can be learned and taught or otherwise transmitted to others has the potential to evolve through this process.
Have other species evolved through cultural evolution?
Cultural evolution, as a process distinctly different from Darwinian evolution, is recognized and accepted by virtually all scientists; but it is generally believed to have had a significant effect only on our own species. Some studies suggest that cultural evolution may also occur among other, non-human primates (Biro et al. 2003; Whiten 2007; Furlong et al. 2008), but clearly it has not had nearly as much impact on these species. And cultural evolution is considered even rarer among other vertebrates, if it exists at all.[14]
However, the embodied view suggests that a revision in this thinking may be necessary. Recall the earlier discussion in which I claimed that the experience of space and time, by all organisms, is an intersubjective process, with some similarities to language. If this thesis is correct, then the evolution of all experience or perception of organisms may very well, just like human language, involve processes other than natural selection. To re-emphasize, random variation and natural selection would play a critical role in creating the kinds of organisms that can interact in societies. But the phenomena emerging from these societies, just like human language, could have a life of their own. They may continue to evolve through intersubjective processes that are basically independent of the genetic makeup of the organisms.
This suggests that cultural evolution, or a process somewhat like it, may have occurred throughout the animal kingdom, just like Darwinian evolution. This doesn't mean that other organisms, even simple invertebrates, learn most of their behavior, in the way that we learn language and myriad other new forms of behavior through our associations with others. Though even simple invertebrates are capable of some learning (Hauser 2000; Griffin 2001), most of their behavior is hard-wired into the nervous system. But the fact that much of it is hard-wired doesn't mean that this behavior must be localized entirely to the brain. It could, and according to the embodied view does, involve interactions with the environment, particularly with other members of the organism's own species. Since these interactions cannot in their entirety be stored or coded for in the genome of an individual organism, the full behavior cannot have a genetic basis, and therefore its evolution arguably cannot be completely explained by Darwinian processes.
Even defined in this way, the notion that cultural evolution occurs throughout the animal kingdom is obviously a very radical idea. It depends on acceptance of the embodied view of perception, which is currently held only by a small minority of scientists. However, even if one dismisses the embodied view entirely, and with it, any talk of cultural evolution occurring among lower organisms, a process somewhat analogous to cultural evolution could have made a major contribution to change long before the emergence of our own species. To understand this, we must look more closely at how cultural evolution occurs, as it acts upon our own species.
What is cultural evolution?
Cultural evolution, like Darwinian evolution, involves variation, in the form of new behavior—language, ideas, images, activities—of all kinds, which is subject to a selection process. Some of it is propagated throughout much or all of the population, while some of it essentially dies out.
Another important parallel or similarity between the two processes-at least in the view of some[15]—is that both involve an informational unit, which is expressed in the form of a phenotype. In Darwinian evolution, of course, the informational unit is the gene. Richard Dawkins (1976) coined the word meme as the unit of cultural evolution, and it was his intention, I believe, that this would be analogous to the gene. But in fact, the way that most people use the term, it corresponds not to the genotype, but the phenotype. Memes are almost always applied to certain forms of human behavior that spread among a population, rather than to the pattern of brain activity that is associated with them.
Despite these similarities, cultural evolution differs from Darwinian evolution in several key respects. First, transmission can occur horizontally, between members of the same generation, as well as vertically, between members of different generations. Second, memes, or the informational patterns that mediate them, do not (necessarily) increase the reproductive fitness of their host organisms. While this certainly may occur, the fitness of memes is determined primarily by their ability to reproduce themselves directly. In fact, this is necessary if transmission is to occur horizontally, where reproduction of individuals is not involved.
These two differences between Darwinian and cultural evolution are widely understood, but there are two additional differences that I believe are not as well appreciated, and which are crucial to our understanding of the latter process. In Darwinian evolution, selection acts on individual organisms, by ensuring that those organisms with a certain phenotype reproduce more than organisms with some other phenotype. In cultural evolution, in contrast, what is selected is (or includes) an interaction between organisms. Though a meme may be a form of behavior performed by an individual organism, in order for it to reproduce itself and therefore be selected, this individual organism must interact with other individual organisms. Memes thus promote socialization; they are the glue that holds societies together.
Finally, there is a two-way interaction between information and expression in cultural evolution. The pattern of brain activity that carries the information mediates—we could say, analogous to the genetic case, codes for—a certain pattern of behavior. This behavior, upon expression by one organism, is then copied by others. But in order to copy this behavior, each of these other organisms must also elicit the corresponding brain activity. So brain activity mediates behavior, but the reverse is also true. Behavior mediates brain activity.[16]
In contrast, the genetic processes that mediate Darwinian evolution are thought to travel in only one direction. According to the Weismann doctrine, the pathway DNA to RNA to protein to phenotype cannot be reversed.[17] There is very little evidence that non-genetic changes in phenotype can result in changes in protein, RNA and ultimately DNA. Indeed, Lamarckian evolution, which holds that characteristics acquired during the lifetime of an organism can be transmitted to their offspring, is largely discredited today precisely because of the believed impossibility of reversing this sequence.[18]
To summarize, cultural evolution is a process in which new phenotypic variations 1) are transmitted horizontally as well as vertically to other organisms; 2) induce expression of both phenotype and informational unit in these other organisms; and 3) are selected by increasing the number of interactions between organisms. Notice that except for the word organisms, this definition could potentially apply to any form of life.
Social evolution and the emergence of proto-organisms.
Consider, for example, the evolution of the first multicellular organisms. The initial step in this process had to be the physical association of individual cells into cell aggregates or colonies. From what we know about cells in the body today, such associations were almost certainly mediated by specific proteins on their surface membranes. If a protein on the membrane of one cell recognized a protein on the membrane of another, the two cells could bind, or attach themselves, together.
While evolution of such recognition molecules could have occurred by Darwinian processes, it would have been difficult. Suppose one cell appears with a mutation resulting in a specific protein capable of binding another kind of protein on another cell. It has to wait for another cell to develop another mutation that creates that protein. Until that happens, the first cell has no selective advantage.
In fact, the situation is even worse than that. It's unlikely that two cells physically associating would have any selective advantage at all. Their mobility would be decreased, while any advantages to be obtained from cooperation—sharing of resources, specialization for different functions, protection from predators—would likely require the presence of many more members of the community.
Suppose, on the other hand, the protein on one cell was capable of inducing a change in a protein on another cell, so that the two proteins could interact. The two cells could now physically associate with each other. Suppose further that the second cell could then induce a similar change in a third cell, which could induce the same change in a fourth cell, and so on. Further suppose that all these changes became inheritable. That is, when the cells with these modified surface proteins divided, they produced daughter cells with the same changes.
Sound far-fetched? In fact, all these molecular processes are known to occur. There is a class of proteins called prions, that can exist in two different shapes or conformations (Prusiner 1998). One of these conformations has the normal biological activity for this protein; for example, it may act as an enzyme. The other conformation is functionally inactive, but it has another, very unusual property. When it comes into contact with another protein of the same kind, it can induce the inactive conformation in it. This converted protein can then induce the inactive conformation in a third protein, and so on. The result is a chain reaction, in which active enzyme molecules are all converted to inactive molecules.
Prions are known to cause several serious diseases in the brain (Safar and Prusiner 1998), by converting essential enzymes into inactive proteins. But they are also found in one-celled organisms such as yeast (Wickner et al. 1999; Serio and Lindquist 2000) and fungi (Coustou et al. 1997; Wickner et al. 1999), where they may be inherited non-genetically. A similar phenomenon—non-genetic inheritance of a novel protein conformation—was reported nearly a half century ago in that familiar staple of high school biology classes, Paramecium (Beisson and Sonneborn 1965).
Prion-like proteins therefore are potentially capable of rapidly transforming a population of single cells, so that all contain a protein that can interact with similar proteins on other cells. This process fulfills the general definition of cultural evolution that I described above. A new phenotypic variation—the ability of a protein to bind to other, similar protein—is transmitted both horizontally, to other cells, and vertically, to daughter cells. The binding process induces the informational unit—an inactive form of the protein—as well as the ability to bind, in these cells. What is selected is not simply the variant protein and its ability to bind, but interactions between cells that have this variant.
The scheme I have just sketched is speculative, of course. There is no direct evidence that this process occurred during the evolution of multicellular organisms, and given the difficulty of obtaining evidence for any events that occurred in this very distant, mostly pre-fossil, past, it is unlikely that any evidence will be forthcoming. But it seems to me highly plausible that something resembling this process occurred. At the very least, the existence of prions and prion-like proteins in both unicellular and multicellular organisms today demonstrates that while cultural evolution is generally thought of as a new process that emerged only with primates, and mostly only in our own species, it reflects a deeper evolutionary strategy that could have been used at a much earlier time.
And as the preceding discussion should have made clear, this kind of evolutionary strategy is much more capable of creating new, social interactions among holons, be they cells or organisms, than Darwinian processes are. A general lesson here is that the Darwinian dynamic, that begins with a novel variation in a single individual, is not very well suited to the evolution of social organizations, which usually entail many individuals with some common features that allow them to interact. While a highly favorable variant may spread rapidly through a population within a few generations, changes that allow social interactions require that several, preferably many, such variants emerge simultaneously. Cultural evolution is uniquely suited to achieving this.
We would not ordinarily refer to processes involving individual cells as cultural evolution, of course. We can define a more general process, though, social evolution, which would include these processes as well as our own cultural evolution, distinguishing them from genetic or Darwinian evolution. Social evolution does not necessarily imply a culture, in the sense of a learned body of information. It only implies that information is transmitted through social interactions, and not (always or entirely) through selection of individual organisms.
When evolution has no choice
There are still other evolutionary processes involved in socialization, however, that may not involve any selection at all. One of the most important of these processes was discovered more than a decade ago, not by evolutionary scientists, but by theorists interested in the properties of networks. Networks are formed whenever large number of units of some kind interact. They include societies of living things, such as molecular and cellular networks, and societies of organisms, but also networks formed by computers and other forms of communication technology.
For decades, scientists had been attempting to model complex natural phenomena with networks based on relatively simple mathematical assumptions. One of these assumptions was that the networks were random, that is, the probability that one unit or node in the network communicated with, or was linked to, another unit was considered no more or less likely than any other link. Towards the end of the millenium, aided by more powerful computers, theorists discovered that many of the most important natural networks are not random, but rather have what is called a small world organization. Most members have relatively few interactions with other members, while a few members have a large number of links (Watts and Strogatz 1998). This feature is referred to as small world, because any individual member, by connecting to a rare highly linked member, is connected in relatively few steps to most of the members of the entire society. This idea actually goes back to the late 1960s, when a famous experiment by Stanley Milgram (1967) suggested that everyone in a large society, even the entire world, was connected by just six links.
Small world organization, particularly a class of network called scale-free, has since been shown to characterize many types of human social systems (Barabasi 2002). But it is also found in the brains of humans and other mammalian species (Hilgetag et al. 2000; Sporns and Zwi 2004; Achard et al. 2006), and still further, in the metabolic networks of cells (Jeong et al. 2000; Wuchty 2001; Yook et al. 2004). While it may possess selective advantagesnot only does it provide highly efficient communication among members, but it is highly resistant to disruption by random loss of a few members (Albert et al. 2000)it is known that such networks always form when a few simple conditions are met. These are primarily a) the network or society is constantly growing, adding new members; and b) new members are more likely to form links with current members that are already highly linked to other members (Barabasi 2002). Anyone who has ever put up an internet site will instantly understand why so much of the internet has a scale-free organization
In other words, the socialization process, at a certain point, occurs not because, or only because, of any selection process, but because certain common conditions determine certain kinds of organization. Evolutionists refer to such changes as constraint-driven processes. The notion that they could be a major contributing factor to evolution is not popular with most scientists. And to be sure, scale-free networks seem to emerge only at a very high level of socialization. Metabolic networks within cells, neuronal networks within brains, and human societies are all exceedingly complex, indeed, one could say they represent the pinnacle of complexity at their particular level of existence. They are the most complex associations of molecules, cells and organisms, respectively, known.
But other kinds of constraint-driven processes could have been involved in the evolution of simpler societies. The emergence of the first simple molecules on earth, such as water, carbon dioxide and urea, was a constraint-driven process. Given certain conditions, carbon, oxygen, hydrogen and nitrogen atoms will always form the chemical bonds that create these molecules. The creation of larger, more complex molecules, such as amino acids, nucleotides and sugars, was also to some extent constraint-driven, as was the formation of primitive macromolecules such as peptides and nucleic acids.
Beyond that, complexity theorist Stuart Kauffman (1993) has demonstrated that the earliest metabolic networks could have resulted from the spontaneous emergence of closed chains of enzymatic catalysis involving just a few dozen molecules. Daniel McShea (2005), who has made a career out of analyzing how and where complexity increases during evolution, has argued that the tendency of the internal components of an organism to become more different over time could constrain the process of natural selection. While McShea's analysis was applied just to individual organisms, since it involves interaction of component parts, it has obvious implications for socialization processes.
Still other evolutionary processes have been proposed (Jablonka and Lamb 2005), some of which are very relevant to the formation of societies. I will not discuss these here, but just conclude by emphasizing that there is potentially far more to evolutionary theory than simply Darwinism. I think critics of the theory who believe that Darwinism is inadequate to account for much of evolution should acquaint themselves with some of this work before jumping to the conclusion that the creation of life on earth required processes or forces for which there is no scientific evidence.
The Three Phases of Evolution
Darwin's theory was never intended to explain all evolutionary history, but only how different species of multicellular organisms emerged. As important as this period was, it comprises only a small fraction of all the evolutionary events on earth. Scientists are of course well aware of this, and seek explanations for how life evolved prior to the emergence of organisms, hundreds of millions of years ago. But because of the absence of fossil evidence, such theories are necessarily highly speculative.
Yet we do have a very general picture of the major events that occurred during this prebiotic era, which have implications for our understanding of evolution that go far beyond this period. We know that the evolution of life must have begun with the formation of macromolecules, sooner or later including DNA, RNA and proteins, that eventually were able to associate into cell-like structures. We know that after the first cells appeared, they could have and almost certainly did evolve into different species in much the same way that organisms do—by random variations in their genetic material, followed by natural selection of the fittest resulting mutant cells. We know that after a great variety of cells were created, some of them began associating into colonies or societies, which took on the properties of quasi-organisms. Some such examples survive today, such as Volvox, a loose association of photosynthetic cells, and Dictoystelium, the slime mold, formed by a reversible aggregation of amoeboid cells.. And we know that further evolution resulted in genuine if still rudimentary organisms, after which the Darwinian story begins.
Translation.
These events, general and fuzzily understood as they are, paint a picture of evolution occurring in three general phases. The first phase is diversification, in which a holon or life form—macromolecules originally, then much later, cells, and still later, organisms—proliferates into a great variety of types or species. This type of evolution, which is more or less what Wilber refers to as translation, involves little or no increase in complexity, and, at least in the case of cells and organisms, appears to be adequately explained by random variation and natural selection. This is a highly competitive period, as the fittest individual holons are selected from probably vaster larger numbers that don't survive.
Transformation.
The second phase, which I often refer to as transformation, is marked by increasing cooperation among individual holons, resulting in socialization and formation of hierarchies, as discussed earlier. Thus macromolecules associated into primitive metabolic networks, composed of dozens or hundreds of such molecules interacting through such processes as enzyme catalysis. Cells associated into colonies or societies, simple at first, but later differentiating into subgroups containing cells specializing in some particular function. And organisms also associate into societies, which have been fairly simple until the emergence of primates, and of course, our species in particular.
Though socialization can be distinguished from diversification, the two processes sometimes strongly interact. Recall that the first step in the complexification of societies is the formation of subgroups, composed of members that perform different functions and which can distinguish members of their own subgroup from members of a different subgroup. This is a diversification process, but when it occurs within a larger unit, in this case a society, it also contributes to both socialization and hierarchical development. The proposal by McShea (2005) for a constraint-driven process in evolution involves this kind of process, except that here the subgroups are tissues or organs within an organism, while the organism is like the entire society. But the principle is the same. Holons or groups of holons that diversify can produce more complexity, if all of them interact within a larger society or other form of organization.
Transcendence: zero-dimensionality.
Finally, the third phase is unification or transcendence. This is arguably the most important evolutionary transition of all, for it produces not simply a new life form, but a new kind of life form, one that, in effect, incorporates all the kinds of life forms that preceded it. Transcendence is a rare, but recurring evolutionary process; it first occurred with the evolution of cells, then again, with the evolution of organisms. As I have discussed elsewhere, both of these forms of life are distinguished from social organizations by 1) their ability to reproduce themselves; 2) their ability to exist outside of individual holons; and 3) possession of what I call a mixed hierarchical structure, distinct from the pure hierarchy or holarchy that characterizes societies (Smith 2009).
Why does transcendence occur? Not much is really known by the process, but in very general terms, we can say that it probably becomes favored when societies become very large and complex. At a certain point, we may speculate, it becomes too difficult for semi-independent holons to cooperate with massive numbers of each other unless some form of centralized control emerges. Cells exert centralized control on molecular networks; organisms impose control on cell assemblies. While non-Darwinian processes are very likely involved, the new form of life would almost certain possess significant fitness advantages.
One of the first and most basic forms of control imposed during evolution of both cells and organisms was limits on growth, on the number of interacting holons. Any cell and any organism has a size limit, beyond which it can't grow, which puts a limit on the number of molecules or cells, respectively, that it can contain. Further growth occurs by expanding not in size, but number—in other words, by reproducing, creating more cells or organisms of limited size. At this point, the cycle begins anew, with diversification of the new life form, and eventually, socialization.
By controlling the growth of its component individual and social holons, a unified holon such as a cell or an organism brings to an end, of course, the processes of socialization and formation of hierarchies. This means that, in one sense, no increase in dimensionality occurs during the transcendence process. In fact, dimensionality at this point starts all over. A newly formed cell or organism has a dimensionality of zero—meaning that it makes no distinctions at all between itself and its environment. However, this zero-dimensionality is, in a very important sense, higher than the dimensionality of any of the social holons that preceded it, since it transcends and includes all of them.
What exactly is this state of zero dimensionality like? One example is provided by extremely simple multicellular forms of life, such as plants, that interact with their environment passively. Plants (and to a large extent, very simple invertebrates) have no sense organs, and generally no capability of independent movement. They are unified mainly in the sense that all their component cells behave in ways that further the survival of the whole plant; their ability to divide, to access nutrients, and to interact with other cells is tightly controlled. While it is difficult for most of us to think of a plant as conscious, we can say that in a purely functional sense, its behavior indicates a minimal distinction between itself and its environment. Like all living things, plants have evolved in such a way as to take advantage of certain aspects of their environment, but they have very little ability to respond to any changes in their surroundings.
It's probably difficult for any of us to imagine the life of a plant. But a state approximating zero-dimensionality is also realized, in a temporary fashion, by many organisms at birth, including our own species. Newborn babies, while having a brain, and a rapidly developing ability both to sense the world and respond to it, are extremely poor at making distinctions between themselves and their environment. While they may have some sense of being a self in a world of others, this sense is vastly less developed than that of any adult. It takes human children a year or two just to learn to experience fully three dimensions of space (Piaget 1973), and our sense of time probably continues to develop in some ways until adulthood, if not longer.
And most important, the process of human development requires socialization. A critical part of the maturation process of human beings is becoming members of increasingly more complex societies—first the family, then the community, then some large system such as a nation, and today, through various mass communication systems, the entire human race. By interacting with others in these increasingly more complex social organizations, each individual, as explained earlier, becomes more complex himself.
Consciousness
If there is one human feature that evolutionary speculation focuses on the most, it's surely consciousness. Could evolution have taken a different path, such that conscious creatures like ourselves never appeared? Or would natural selection, along with other evolutionary processes discussed here, strongly favor consciousness?
The notion that increasing consciousness is a general evolutionary trend is supported by noting that consciousness appears to be closely correlated with other trends discussed here. Certainly that is the case with our species. I emphasized earlier that, by anyone's definition, the human brain is more complex than the brains of other organisms.
We are also the most highly socialized organism, and as discussed elsewhere (Smith 2009), capable of the highest degree of dimensionality in our experience of space and time.
We don't know, of course, how many other species are conscious, and to what degree. But ask any scientist, or non-scientist for that matter, which animals are or might be conscious, and the list will begin with the higher vertebrates, with the most complex brains other than our own, and stop at some point among lower vertebrates or perhaps invertebrates. Everyone, in other words, makes the assumption that larger, more complex brains are closely correlated with greater consciousness.
There is a philosophy of mind, called functionalism, that could be said to embody this notion (Dennett and Hofstadter 1981; Dennett 1991). Functionalists claim that consciousness emerges from the interactions of information processing units, regardless of their nature. In this view, consciousness does not have to be restricted to a living organism. Computers could and would be conscious if they have a pattern of organization and activity identical to that of the brain of a conscious organism. Functionalism thus makes a powerful argument that the socialization process, applied to almost any kind of holon, might have inevitably resulted in consciousness.
While functionalism has some strong critics (Searle 1992; Chalmers 1996; Griffin 1998; Seager 1999), very few if any of them would deny that organization is paramount. Philosopher John Searle (1992), for example, apparently believes that only organisms with brains composed of neurons can be conscious, but he doesn't deny that whether something is conscious, or how conscious it becomes, must depend on the number of these neurons and the complexity of their interactions. In fact, it is well-established that the main difference between the human brain and that of other species, even including simple invertebrates, is largely the greater interconnectivity of its cells. The nerve cells found in invertebrates are not very different, in their structure, morphology and electrochemical properties, from those in our brain (Bullock 1965). So it could be argued that once excitable cells like neurons appeared, at least hundreds of millions of years ago, large complex brains and consciousness were virtually inevitable.
On the other hand, there has always been one very problematic aspect of discussions of the evolution of consciousness: it isn't clear that it provides any fitness advantage to organisms. At the root of this argument (and several others in philosophy of mind) is a purely hypothetical being that philosophers refer to as a zombie; it is physically identical to some human being[19] yet has no consciousness (Chalmers 1996; Seager 1999). The question resolves around whether zombies are possible, or conceivable. Not all philosophers believe they are, but if they are, it is very difficult to see how consciousness provided organisms with any survival advantage. If organisms, including human beings, lacking consciousness could do everything the conscious organism could do, why would they not evolve and be selected, without the superfluous burden of consciousness? Indeed, as I discussed earlier, according to the standard scientific worldview, consciousness is supposed to be completely determined by some pattern of activity in the brain (Chalmers 2000). But according to this view, so is all of our behavior; it is all the result of physiological processes in the brain. So what is the evolutionary purpose (i.e, fitness advantage) of consciousness?
One solution to this problem is to regard consciousness as an inherent property of all matter, a fundamental feature, like mass or charge, that is simply accepted without further explanation. This move, usually part of a philosophical view known as property dualism (Chalmers 1996; Seager 1999), has the additional advantage of solving (or perhaps more accurately, avoiding) the materialist dilemma of how consciousness, seemingly so different from physical processes, can emerge from them. Of course, it avoids that question by raising another one: what exactly does it mean to say that all matter is conscious? Very few scientists or philosophers are comfortable with even contemplating this possibility. Some have claimed that there is in fact experimental support for concluding that consciousness only emerges in fairly complex brains (Ramachandran and Hirstein 1997).
I am not going to argue strongly for this panpsychist view of consciousness here (see Smith 2009). But if we do adopt it, then the fact that anything is conscious does not have to be explained by evolution, any more than the fact that anything is composed of atoms. What does have to be explained is why consciousness increases. And we now have our answer in the evidence for the other evolutionary trends discussed previously. To the extent that consciousness is closely associated with forms of matter and life that have increased in complexity, we would expect that consciousness would also increase in complexity.
Our Evolutionary Future
I have argued here that there are certain very large-scale evolutionary trends that have recurred several or more times over the history of the earth. These include increasing complexity, socialization, hierarchy, dimensionality and consciousness, played out over periods of alternating diversification and unification. If we ask ourselves how these trends might extend into the future, one phenomenon seems to stand out as the possible next step: a higher state of consciousness. I am not going to discuss here all the evidence for this state. But if we accept the state as realand I will have a bit more to say about what that means in a momentcould it not be understood as the experience of a new form of life, more complex than any before it, resulting from another great unification process? A form of life encompassing the entire earth, and particularly all human societies?
This is not the only way to understand higher consciousness, of course. But for those who do recognize it as a genuine phenomenon that requires explanationsome kind of connection with the rest of our body of knowledge—I think there are only two other possibilities worth serious consideration: 1) it is completely independent and prior to all forms of life on earth, in other words, not associated with any particular physical system; or 2) it is a product of the individual human brain just as—in the mainstream scientific view—our ordinary consciousness is.
The mystical view of higher consciousness.
The first alternative is the classic or traditional mystic view, and frequently espoused by Wilber.[20] Wilber, like several others before him, perhaps most notably Sri Aurobindo (1985), embraces evolution, and seeks to explain it by appealing to higher consciousness. In his view, higher consciousness is understood as a guiding force, or eros, that provides not only a direction but a goal and purpose for evolution on earth. David Lane has pointed out that Wilber's evolutionary views do not simply supplement scientific theory, which I regard as a valid undertaking, but in several key places openly contradict it. Or perhaps more precisely, Wilber seeks to use the eros concept in places where evidence strongly suggests it's not needed.
However, it seems to me that there is a more fundamental problem with the mystical worldview. It's a form of dualism, with all the well-known difficulties associated with that view. On the one hand, consciousness is said to be independent and prior to the physical world; on the other, it creates this world, and in the view of some, like Wilber, guides its evolution. This implies some kind of interaction with it. How can both of these statements be true?[21]
One possible solution to this problem is offered by the advaita or nondual philosophy found in the teachings of such twentieth century gurus as Sri Ramana Maharshi, Nityananda, and Nisargadatta (Frydman 1999; Hatengdi and Chetanananda 2001; Powell 2006). Nondualists believe in a single reality that most of us are unable to perceive or experience because we are in a state of sleep, dreaming or illusion. In this view, the entire physical world, including everything known to science, is part of this illusion. Since it is all unreal, there is no distinction between higher consciousness and anything else. There is only a single reality (or more precisely, there is neither one or other). For this reason, many non-dualists, I think, are not inclined to criticize or reject the scientific worldview, including evolutionary theory, any more than they are inclined to reject the apparent evidence of our senses in everyday life. The idea is not to explain illusory events, but to awaken from them.[22]
One might succinctly describe the nondual view, therefore, as separation of church and state. We won't criticize your explanation of the world, say the nondualists; just don't attempt to use your science to explain awakening. But such separation, ironically, reeks of dual, not nondual. As much as the nondualist insists that ordinary consciousness is not real, and therefore not part of a dualist system, it seems to me that this distinction continually returns to haunt him.
Consider an obvious analogy or metaphor for the relationship between illusion and awakening: that of the ordinary dreaming that we all do. When we awaken, we don't attempt to explain how or why certain things happened in the dream, because we recognize that none of it was real. From the perspective of the dreamer, these events might have had causes (at least in our more coherent dreams), but from the perspective of being awake, the question doesn't really matter. Since the dream is a temporary, make-believe world, explanations are beside the point. So far, so good.
The problem with this metaphor (and with other popular metaphors, such as virtual reality, the events on a movie screen, certain optical illusions, or even characters in a novel) is that dreams do have an aspect of reality. While the events of dreams may not occur in what we regard as the real world, the experience of engaging in these events has real, physiological causes. Real world events occurring inside our brainsneural activity in certain well described regions—are considered by scientists to be the causes of dreams, and it is assumed that any dream experience can be ultimately explained in terms of some specific pattern of this activity.
The nondualist might reply that dreaming is an imperfect metaphor for the relationship between illusion and awakening into higher consciousness. I'm sure it is. But it still illustrates the fundamental problem facing the nondualist: illusion and awakening are two different states. Even if the events that we experience during ordinary consciousness are unreal, this consciousness is still a state, and quite apparently distinct from the state of real awakening. One only has to ask questions such as: Why do we experience illusion? or What is the difference between someone who is asleep and someone who is awake? to see the problem. It's also illustrated by the very existence of methods or techniques that are claimed to help an individual awaken.
It's not my intention to denigrate the nondualist view, which I personally find very powerful and in many respects persuasive. In fact, the dream metaphor is widely held by mystics, including many who might not regard themselves as living within a nondualist tradition. My only point is that there is abundant evidence for an interaction of higher consciousness with our ordinary state; the two are neither completely separate, nor completely understandable as a single state of reality. Perhaps the most obvious evidence of this is the effect that an experience of higher consciousness has on someone. She thinks about it, talks about it, perhaps theorizes about it. All of this activity involves physical events in the brain, events that we must conclude were caused by the experience of higher consciousness. If there had been no experience of higher consciousness, these events would not have occurred.[23]
This being the case, I think it's relevant to bring science into the picture. A mystic could argue that, just as a scientific explanation of dreaming lies totally beyond the comprehension of the dreamer, so the explanation or understanding of illusion lies totally beyond ordinary consciousness. I think that's a very good point; it may well prove to be the case. But to the extent that the two states interact, we can at least ask how these interactions impact the world that science is capable of investigating.
Is higher consciousness mediated by events in the human brain?
This is exactly the approach taken by those in the second camp, who believe that higher consciousness is the result of physiological processes in an individual's brain. This view is held, explicitly or implicitly, by most researchers who are actively seeking to identify these physiological processes, or correlates. The earliest of these studies, several decades ago, used EEG recordings, but more recently this research has been supplemented with neuroimaging technology, such as functional magnetic resonance (Orme-Johnson and Haynes 1981; Newberg 2002; Lazar et al. 2005; Holzel et al. 2007; Lutz et al. 2008). The guiding assumption underlying most of this research is that the brains of individuals realizing higher consciousness must be different in some way from that of subjects who are not. And it would seem very difficult to argue otherwise, for as we have just seen, experience of higher consciousness does have very apparent effects on the individual brain.
On the other hand, there are several problems with this view.[24] One of them echoes the dilemma of localizing ordinary consciousness to the brain: what function does it serve that would allow it to be selected for? Does experiencing higher consciousness promote survival or well-being in some way? Possibly, but if higher consciousness is mediated by activity in the brain—which, to re-emphasize, is the guiding assumption of most of these scientific studies of it—why isn't that activity itself sufficient? Survival involves functional behavior in the environment, and the zombie argument holds that consciousness is unnecessary for this. Even if one wanted to argue that experience of higher consciousness has internal effects—say, promoting health, harmony and longevity among the various physiological processes of the body—these effects still, according to science, must be mediated by neural activity. Again, consciousness seems superfluous.
In the case of higher consciousness, though, the problem of selection is even worse than that. It seems that some people, perhaps very few, perhaps more, are capable of experiencing higher consciousness. It does not seem obvious that they inherited this ability in the form of a rare variation, so where did it come from? Are they simply making use of parts of the brain that all or many of us have? If that's the case, these parts have gone unused in large numbers of people for a very long time. So why did they evolve in the first place? Are they spandrels, a term coined by Gould and Richard Lewontin (1979) for side effects of evolutionary adaptations that may later turn out to have useful functions? Many evolutionists contest the very existence of spandrels in this sense. In any case, it's difficult for me to conceive of the capacity for having such a profound experience as purely a side effect, a consequence of evolutionary processes that were selected for an entirely different reason.
Another problem with the view that higher consciousness is mediated entirely by processes in the individual brain should be apparent from the characteristics of this state. Virtually everyone who has experienced it has described it as beyond all thought, feeling and behavior. Another common feature is the experience of a unity or oneness with something far grander than the individual, often described as the entire world or even the entire universe.
Such experiences might be of something that is not real—in other words, the experience is just an illusion—and probably many skeptical scientists are inclined to view them in this way. But that simply throws us back to the question of how such illusions could have evolved. There are many well known illusions, particularly visual ones, that the human brain is capable of generating, but they generally have good evolutionary explanations. Perhaps someone will come up with an analogous explanation for how experience of higher consciousness could be an illusion, but I have yet to see it.[25]
It seems to me, therefore, that the experience most likely does correspond to something real. If that is indeed the case, it's difficult to see how or why it would be mediated by an individual brain. In our ordinary state of consciousness, we have no direct experience of anything beyond ourselves and the immediate environment; and as far as we know, every other organism on earth faces the same limitations. If higher consciousness is the experience of a much larger and more complex form of life, wouldn't it be very closely associated with this form of life?
As I pointed out earlier, when new life forms evolved in the past that transcended all classes of existence that preceded them—first, the cell, later the organism—they were characterized by zero-dimensional experience. The defining feature of this state is the inability to make any distinction between self and other. This, of course, is also a defining feature of higher consciousness. So this experience is not only of something that would seem to transcend our ordinary existence, but it is the kind of experience that has characterized other evolutionary periods of transcendence.
In conclusion, while there may be correlates of higher consciousness in the human brain, I think it's a mistake to think that identifying these correlates will tell us very much about the experience. The embodied view I have discussed here implies that even our ordinary consciousness is not localized entirely to the brain; but the individual brain is certainly a major player in the embodied dynamic. In the case of higher consciousness, correlates in the brain are very likely only the faintest signals of a consciousness associated with a much higher form of life.
The end of evolution?
Higher consciousness, by its very nature, is likely to remain forever beyond any full scientific explanation. But it seems to me the best way to understand it, at our current level of scientific knowledge, is as the manifestation of an evolving new form of life, one that stands to individual humans and their societies much as cells stand to molecules, and organisms to cells. This view may seem radical to a scientist, but it shouldn't be. If one can believe that our ordinary consciousness results from the activity of billions of neurons in the brain, why can't one believe that a much higher form of consciousness emerges from the interactions of billions of far more complex individuals (and uncountable numbers of other forms of life) on earth?
This conclusion does not necessarily imply that everyone, or even more than a few individuals, will awaken to become this higher form of life. Indeed, if earlier evolution is a guide—and obviously, I have been using it as a guide throughout this discussion of evolutionary trends—complex societies will be a critical and integral form of any higher life. In the future, these societies will likely be composed of either human beings, and/or forms of human-invented artificial life, probably including hybrids of computers with humans. Whatever their composition, though, I believe the growth of these societies will become highly constrained—an effect that most of us, in the developed world, already see as a necessity. We have seen that when cells evolved, they placed limits on the number of molecules they were composed of; and that when organisms evolved, they placed limits on the number of cells they were composed of. It seems to me that any putative higher form of life will face the same problem with human societies, and will solve it in basically the same manner.
In fact, this evolutionary view, for whatever it's worth, strongly suggests that we are heading towards the end of evolution—on earth, that is. Just as evolution of molecules largely ended with the emergence of cells, and evolution of cells eventually ended following the emergence of organisms, I believe the evolution of a genuinely higher, trans-human form of life would indicate that the days of evolution of new species of organisms are numbered. Our species may continue to evolve, not through natural selection but through cultural evolution and through increasing integration with computers, but as we obtain increasingly greater knowledge of our environment, I think the day will come when further evolution becomes superfluous. Further evolution would and could occur, in this view, only if the earth life form could reproduce itself, presumably by colonizing other planets.
All of this is sheer speculation, of course, simply a prediction of future trends based on those of the past. Evolution again and again has proved to have astonishing creativity, unleashing all manner of things that we humans—if we could have lived back in those days—would never have predicted. The general trends that I have outlined here, though, are broad enough, I think, to encompass an enormous amount of novelty, while still giving us some sense of where we are going.
ENDNOTES
* See: David Lane, "Frisky Dirt: Why Ken Wilber's New Creationism is Pseudo-Science"; David Lane and Andrea Diem-Lane, "Random Mutations
in Molecular Biology: Why Ken Wilber's Creationist Hummer Got Recalled" and "The Faith of Physical Causes: Presenting the Evidence for Biological Evolution", www.integralworld.net
[1] Tononi et al. (1994) developed a mathematically precise definition of complexity that they applied specifically to the brain. It aims to recognize the importance of both different, or independent, patterns of brain activity as well as their integration into overall brain function. Their definition can be considered a much more specific version of the general one that I am using here.
[2] See Table 1 in Summary Statistics at www.iucnredlist.org
[3] This is a theoretical maximum for the number of states, assuming that every possible state of any individual component can combine with every possible state of any other individual component. In reality, there are likely to be constraints; for example, In real systems beyond a certain size, individual components generally cannot interact with all other components, which will reduce their complexity. In fact the maximum number of possible states of a system will usually be considerably less than this theoretical number.
Still even with such constraints, a disproportionate increase in states occurs as the number of components increases. Moreover, complexity may increase if the connections between components may vary in such parameters as strength and valence (i.e., positive or negative effect). Individual components may also access some states independently of connections with other components, through, for example, internal changes in their own component holons.
[4] The ability to recognize or distinguish different individuals is closely associated with what I call three-dimensional experience, and for the most part emerged with vertebrates, and some very intelligent and advanced invertebrates like cephalopods. However, some examples have been documented among arthropods. Crayfish (Schneider et al. 1999; Gherardi et al. 2005) and crickets (Delago and Aonuma 2006; Iwasaki et al. 2006), recognize and avoid individuals that they have previously lost to in aggressive encounters, though it's possible that such distinctions could be made on the basis of classes rather than individuals. Two social insect species, one a paper wasp (Tibbetts 2002) and the other an ant queen (Dreier et al. 2007) have also been shown to distinguish different individuals. Honeybees can actually be trained to recognize different human faces (Dyer et al. 2005).
[5] Even so-called solitary species of arthropods have the ability to make class distinctions, distinguishing kin from non-kin (Metzger et al. 2010), and male from female (Askari and Alishah 1979; Godfray 1994), using the same kinds of processes that social insects use. Thus it is not necessary to live in constant physical contact with other members of the same species to be a member of a society. In fact, in these and other so-called asocial organisms, vertebrates as well as invertebrates, the society may include not simply members of the same species, but members of other species, in other words, an entire eco-system. The organism is engaging in social interactions with other species that can be just as complex as those that social organisms exhibit within colonies or societies.
[6] The ability of social insects to distinguish caste memberships is based on their ability to distinguish complex mixtures of chemicals, known as pheromones, that are associated with these insects, and which act as membership tags. This is the fundamental class distinction that these organisms make, with other distinctions, such as those among different caste members, generally deriving from this. But many arthropods can make other class distinctions, in such modalities as light (different colors; Briscoe and Chittka 2001; Van der Velden et al. 2008) and sound (different patterns of intensities and other qualities; Hoy 1992; Fullard and Yack 1993).
[7] I don't claim this as evidence for a second dimension of time in the universe. See Smith (2009) for further discussion of this.
[8] The work by Bach-y-Rita (1972) is particularly remarkable. Bach-y-Rita worked with blind subjects, developing an apparatus that allowed them to move through their environment. The device consisted of a camera which fed visual information to an array of vibrators on the person's skin (back or thigh were generally used). By sensing the pattern of vibrations, the subjects could obtain information about objects in the environment, much as one could identify letters that are traced out on the skin.
What is extraordinary about this work, though, is that the subjects claimed they could actually see their environmentthat the process of interpreting the pattern of vibrations was visual, not tactile. To the extent that they actually did experience vision, this supports the embodied view. A major thesis of this view is that experience of the environment results from certain dynamic relationships we have with it, rather than activity in some localized part of the brain. In the case of vision, these relationships include such phenomena as objects appearing larger when they are approached, and smaller as we move away from them, as well taking on different shapes when we move around or to the side of them. Such relationships were preserved in Bach-y-Rita's apparatus, and thus in the embodied view, this is why the subjects experienced vision, even though the activity in their brains took place in areas normally mediating touch.
[9] This point can be made independently of the raging debate over whether language is innate—completely hard-wired into the brain—or learned (Pinker 2007). Though there are a few researchers who argue that we are hard-wired to understand a large number of basic concepts (Fodor 1998), the fact remains that the meanings of these concepts, like the meanings of any words, are social. So the real question is how much of the structure of language—grammar, syntax, and so on—is innate, and how much is learned.
[10] Actually, human language can be regarded as more or less equivalent to our experience of space and time. That is, the way we experience our environment—more precisely, the ways in which our experience of the environment differ from that of other organisms—is largely determined by language. For example, we have a much deeper experience of time than other organisms, which is closely associated with our ability to label objects and retain these labels throughout our life. We also experience objects in space differently, because we have concepts about these objects that are an intimate part of our perception of them. My claim here is that to be able to express and understand human language is equivalent to being able to experience the higher dimensions of space and time that are available only to our species.
So when I assert that all experience of space and time by other organisms is intersubjective, I'm basically just saying that they derive their experience in the same way we do—from the complexity of their social interactions. I'm extending the process that we know is in play for our species to all other species.
[11] A notable exception to this is physicist Itzhak Bars, who believes there are four spatial dimensions and two dimensions of time. Bars argues that by incorporating these extra dimensions into our view of reality, many previous problems or paradoxes that emerge in models with a single dimension of time can be solved . See:
http://physics1.usc.edu/%7Ebars/research.html#2T. Brief summaries for the layperson can be found in New Scientist (Oct. 13 2007) and Discover (May 29 2008).
[12] Kaku (1996), p. 231. For an argument against a second dimension of time, see Deutsch (1998), and my response to it in Smith (2009).
[13] Fitness may include properties that promote interactions in certain ways, of course, just as language areas in the brain promote communication between people. But properties that emerge only from such interactions—again, such as the meanings of particular words—cannot evolve through a process that selects individual fitness.
Moreover, even in the case where individual fitness can promote social interactions, such changes normally have to emerge in several organisms at once. A single organism with a language area in its brain would not have any selective advantage over other organisms lacking the language area; on the contrary, the language area only provides greater fitness to an individual if there are other organisms with the language area, that can communicate with that individual. As I will discuss further below, it is thus difficult for Darwinian processes to account for any adaptations that promote greater social interactions, because the changes must occur in several organisms at once.
Finally, I reiterate a point made in note 9, that many scientists believe that other, more basic aspects of language, such as grammar and syntax, may to some extent be learned by each generation, rather than the product of natural selection (Pinker 2007).
[14] This conclusion, though, may depend on precisely how cultural evolution is defined. Jablonka and Lamb (2005) define a process they call social evolution, which involves transmission of behavior through learning, exemplified by parent-offspring transmission in many birds. They distinguish this from human cultural evolution by the fact that language or symbols are not involved. What they call social evolution, however, could, along with cultural evolution, be subsumed under the still broader rubric of social selection, to be discussed below.
[15] Not all researchers in the field of cultural evolution believe that the concept of an informational unit, such as the meme, is useful, or at the least, adequately describes the entire process. For example, a great deal of cultural transmission may result when individuals learn from a variety of sources, not copying a single idea or form of behavior. Moreover, much of this learning may come from trial-and-error, so that there is no real copying process at all (Boyd and Richerson 1985). Nevertheless, I believe the main features of the process as I will describe them are fairly universal.
[16] We now know that a key factor in the reproduction of memes is played by mirror neurons, neuronal networks in the prefrontal cortex that are active not only when an animal performs some specific action, but also when it sees another animal performing the same action (Gallese and Goldman 1998; Rizzolatti and Craighero 2004).
[17] Weismann died in the early twentieth century, long before the discovery of DNA. However, at the time of his death it was recognized that there had to be physiological processes responsible for inheritance, beginning with the reproductive apparatus, and his theory or doctrine stated that there was a one way path leading from these processes to the phenotype. His doctrine was formulated specifically to deny the possibility of Lamarckian evolution, and in the opinion of most scientists, it is still valid today. Though recent discoveries in molecular biology and other areas suggest the possibility of inheritance of acquired characteristics in some cases (Jablonka and Lamb 2005), their interpretation is controversial, and in any case, such cases seem to be fairly uncommon.
[18] However, new discoveries in molecular biology, physiology and behavior are now suggesting, at least to a few scientists, the possibility of Lamarckian evolution under some circumstances. This work is reviewed in detail by Jablonka and Lamb (2005).
[19] Different kinds of zombies have been proposed, which have different philosophical consequences. The kind of greatest interest to philosophers is an exact, atom-by-atom duplicate of a human being, because if such a life form can be conceived that has no consciousness, this strongly suggests that physicalism—which holds that consciousness is determined entirely by physical processes—is false. But the argument against natural selection does not depend on this kind of zombie. It only depends on the possibility of a zombie that is functionally and behaviorally identical to a human being, because (assuming it had genes like ours) this would be exactly as evolutionarily fit as a conscious human being. And as the Stanford Encyclopedia entry on zombies (http://plato.stanford.edu/entries/zombies/), notes: plenty of physicalists accept that merely behavioral or functional duplicates of ourselves might lack qualia [i.e., consciousness].
[20] Wilber, as always, is confusing, and subject to multiple interpretations (see also the following two notes). He has long argued that consciousness must be associated with some physical system, and his famous four-quadrant diagram makes this association explicit. Further, he has expressed in the past support for David Chalmers' (1996) view of property dualism, which claims that consciousness is an inherent property of all matter. But he also talks about a consciousness that is the ground of all existence. I don't see how this latter view can be consistent with property dualism, or with the 4Q view of consciousness as a dimension of holons that also includes physical properties.
[21] This problem of dualism is exacerbated by the fact that some who hold the mystic view, such as Wilber, believe it is also consistent with the second alternative I will discuss here, that there are physiological correlates of higher consciousness in the brain. This latter position was also explicitly held by one of the first mystics to develop the notion that higher consciousness guided evolution, Sri Aurobindo (1985). Aurobindo actually claimed that as individuals realized the highest states of consciousness, there would be changes not only in the brain, but in other parts of the material world, that could be demonstrated scientifically (see Kazlev 2006). So on the one hand, this consciousness is independent of all of the physical world; on the other hand, it can induce changes in it.
[22] I don't claim that all nondualists take this approach. In fact, Wilber at various times has claimed to hold to a nondual view, or at least has said that his system is consistent with such a view. But the important point is that this approach is very consistent with non- dualist teachings. For a highly articulate exposition of this view, see the blog postings of Conrad Goehausen, aka Broken Yogi (www.brokenyogi.blogspot.com). Conrad has a very good understanding of modern science, and to the best of my knowledge is a strong supporter of evolutionary theory. But he also espouses a non-dualist view, and is a strong critic of Wilber's general approach of attempting to connect and/or to find support for such teachings with scientific findings.
[23] The notion that consciousness has an effect on brain processes might seem to contradict the zombie argument that consciousness is superfluous to the relationship between brain and behavior. If some conscious experience results in our thinking or acting in ways that we would not have in the absence of that experience, doesn't that prove that consciousness itself can cause certain thoughts to occur?
Certainly many, probably the great majority of, philosophers would argue that it can. But that is not relevant to the zombie argument. If consciousness itself is the result of processes in the brain, the zombie argument maintains that we can cut out this middleman, so to speak, as the processes themselves are sufficient to cause the thoughts. There is no evidence I'm aware of that consciousness can cause certain thoughts to occur that can't be interpreted in this way. Some have argued that a zombie is capable of having even the thought I am conscious (though exceptions for cases like this could be made, and the zombie argument against evolution of consciousness would still be very strong).
So in the specific case of someone experiencing higher consciousness later thinking about it, the argument would be that the experience of higher consciousness was mediated by certain brain processes, and these processes in and of themselves are sufficient to cause the later thoughts about higher consciousness. This does not affect the argument that higher consciousness in itself is linked in some way to brain processes, regardless of whether or not is mediated by such processes, because the zombie is only a hypothetical construct. All or virtually all philosophers believe, regardless of whether the zombie argument is valid or not, that consciousness actually exists, and likewise, we can accept for the sake of this discussion that higher consciousness also exists.
[24] Aside from conceptual problems, there are also technical ones, which I have discussed in Smith (2009).
[25] There may be aspects of the experience of higher consciousness that appear explainable by illusion. For example, some studies suggest that the experience is associated with regions of the brain involved in our perception of space and time (Newberg et al. 2002). One could argue that activity in these areas inhibits our normal experience of these dimensions, resulting in the well known experience of infinity, or being beyond dimensions entirely (or as I discuss later, having zero-dimensional experience).
The problem with interpreting the state in this way is that this altered experience of space and time is only one feature of higher consciousness. There are many other features, such as transcendence of thought and feeling, that would not result from activity in areas of the brain responsible for experience of space and time. Thus one would have to postulate, then find experimental proof for, activity in all the other relevant regions. Indeed, the very nature of higher consciousness suggests that, if it's an illusion, it's one that has captured all the ordinary functions of the brain. This is very unlike illusions that are well known and studied. And to reiterate, there is no indication of an evolutionary explanation for such illusions.
REFERENCES
Abbott EA (1992). Flatland: A Romance of Many Dimensions (New York: Dover Thrift
Adami, C. (2002) What is complexity? Bioessays 24: 1085-1094.
Achard, S., Salvador, R., Whitcher, B., Suckling, J., Bullmore, E (2006) A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J Neurosci 26, 63-72.
Albert, R, Jeong, H., Barabasi, A.L. (2000) Error and attack tolerance of complex networks. Nature 406: 378-382.
Allen, T.F.H., Starr, T.B. (1982) Hierarchy: Perspectives for Ecological Complexity (Chicago: University of Chicago Press)
Askari, A., Alishah, A. (1979) Courtship behavior and and Evidence for a Sex Pheromone in Diaeretiella rapae (Hymenoptera: Braconidae), the Cabbage Aphid Primary Parasitoid. Ann. Entomol. Soc. Amer. 72, 749-750
Aurobindo S. (1985) The Life Divine (Twin Lakes, WI: Lotus)
Bach-y Rita P. (1972) Brain Mechanisms in Sensory Substitution (New York: Academic Press)
Barabasi, A.L. (2002) Linked (New York: Perseus)
Bargmann, C.I. (2006) Chemosensation in C. elegans. WormBook (Oct. 25): 1-29.
Beisson, J., Sonneborn, T. M. (1965) Cytoplasmic inheritance of the organization of the cell cortex in Paramecium aurelia. Proc. Natl. Acad. Sci. USA 53, 275-282.
Bennett, C.H. (1988) Logical Depth and Physical Complexity in The Universal Turing Machine— a Half-Century Survey, Herken, R., ed. (Oxford: Oxford University Press) pp. 227-257
Bielecki A (1999) The role of eye-like spots of parasitic leeches (Hirudinea, Piscicolidae) in searching for fish-hosts Wiad Parazytol 45: 339-345.
Biro, D., Inoue-Nakamura, N., Tonooka, R., Yamakoshi, G., Sousa, C., Matsuzawa, T. (2003) Cultural innovation and transmission of tool use in wild chimpanzees: evidence from field experiments. Anim Cogn 6, 213-223.
Briscoe, A.D., Chittka, L. (2001) The evolution of color vision in insects. Annu. Rev. Entomol 46: 471-510.
Bounoutas, A., Chalfie, M. (2007) Touch sensitivity in Caenorhabditis elegans Pflugers Arch 454: 691-702.
Boyd, R., Richerson, P. (1985) Culture and the Evolutionary Process (Chicago: University of Chicago Press)
Bullock, T.H. (1965) Structure and Function in the Nervous Systems of Invertebrates (New York: Freeman)
Cavalli-Sforza, L., Feldman, M.W. (1981) Cultural Transmission and Evolution: A Quantitative Approach (Princeton, NJ: Princeton University Press)
Chaitin, G. (1973) Abstracts of Papers, 1973 IEEE International Symposium on Information Theory, June 25-29, 1973, King Saul Hotel, Ashkelon, Israel, IEEE Catalog No. 73
Chalmers, D. (1996) The Conscious Mind (Oxford: Oxford University Press)
Chalmers, D. (2000). What is a neural correlate of consciousness?, in Neural Correlates of Consciousness: Empirical and Conceptual Questions Metzinger, T., Ed. (Cambridge, MA: MIT Press) pp. 18-39.
Changizi, M.A. (2001) Universal laws for hierarchical systems. Comments on Theoretical Biology 6: 25-75.
Cheney D.L, Seyfarth, R.M. (2007) Baboon Metaphysics Chicago: University of Chicago Press.
Cooper, G.M., Brudno, M., Green, E.D., Batzoglou, S., Sidow, A. (2003) Quantitative Estimates of Sequence Divergence for Comparative Analyses of Mammalian Genomes. Genome Res. 13, 813-820
Coustou, V., Deleu, C., Saupe, S., Begueret, J. (1997) The protein product of the het-s heterokaryon incompatibility gene of the fungus Podospora anserina behaves as a prion analog. Proc Nat Acad Sci USA 94, 9773-9778.
Dawkins, R. (1976) The Selfish Gene (Oxford: Oxford University Press)
Dawkins, R., Gould, S.J. (1997) Human Chauvinism Evolution 51, 1015-1020
Deacon, T.W. (1998) The Symbolic Species (New York: W.W. Norton)
Delago, A., Aonuma, H. (2006) Experience-based agonistic behavior in female crickets, Gryllus bimaculatus. Zoolog Sci 23, 775-783.
Depew, D.J., Weber, B.H. (1997) Darwiinism Evolving (Boston, MA: MIT Press)
Dennett, D.C. (1991) Consciousness Explained (New York: Little, Brown)
Dennett, D.C., Hofstadter, D.R., eds. (1981) The Mind's I (New York: Basic)
Deutsch, D. (1998) The Fabric of Reality (New York: Penguin)
Dobzhansky, T. (1973) Nothing in biology makes sense except in the light of evolution Amer. Biol. Teacher 35, 125-129
Dreier, S., van Zweden, J.S., D'Etorre, P. (2007) Long-term memory of individual identity in ant queens. Biol Lett 3, 459-462.
Dunbar, R.I.M. (2003) The social brain: Mind, language, and society in evolutionary perspective. Evolutionary Anthropology 6, 178-190.
Dyer, A.G., Neumeyer, C., Chittka, L. (2005) Honeybee (Apis mellifera) vision can discriminate and recognise images of human faces. J. Exp. Biol. 208 (Pt24), 4709-4714.
Flannery, T. (2005) The Weather Makers. How Man is Changing the Climate and what it means for Life on Earth (New York: Grove/Atlantic)
Fodor, J. (1998) Concepts: Where Cognitive Science Went Wrong (New York: Oxford University Press)
Frydman M, translator, ed. (1999) I Am That: Talks with Sri Nisargadatta Maharaj (Kala Goda, Mumbai, India: Chetana Private, Ltd.)
Fullard, J.H., Yack, J.E. (1993) The evolutionary biology of insect hearing. Trends Ecol Evol. 8, 248252.
Furlong, E.E., Boose, K.J., Boysen, S.T. (2008) Raking it in: the impact of enculturation on chimpanzee tool use. Anim Cogn 11, 83-97.
Gallese, V., Goldman, A. (1998) Mirror neurons and the simulation theory of mind-reading. Trends Cogn Sci 2, 493-501.
Gherardi, F., Tricarico, E., Atema, J. (2005) Unraveling the nature of individual recognition by odor in hermit crabs. J Chem Ecol 31, 2877-2896.
Godfray, H.C.J. (1994) Parasitoids: Behavioral and Evolutionary Biology (Princeton, NJ: Princeton University Press)
Gould, S.J., Lewontin, R.C. (1979) The spandrels of San Marco and the panglossian paradigm: a critique of the adaptationist programme. Proc. Roy. Soc. London B 205, 581-598.
Gould, S.J. (1990) Wonderful Life: The Burgess Shale and the Nature of History (New York: Norton)
Gould, S.J. (1996) Full House: The Spread of Excellence from Plato to Darwin (New York: Harmony)
Gould, S.J. (2002) The Structure of Evolutionary Theory (Cambridge, MA: Harvard University Press)
Griffin, D. (1998) Unsnarling the World Knot: Consciousness, Freedom and the Mind-Body Problem (Berkeley, CA: University of California Press)
Griffin, D.R. (2001) Animal Minds (Chicago: University of Chicago Press)
Hatengdi, M.U., Chetananda,, S. (2001) Nityananda: In Divine Presence (Portland, OR: Rudra.Press)
Hauser, M.D. (2000) Wild Minds (New York: Henry Holt & Co.)
Hilgetag, C.C., Burns, G.A.P.C., O'Neill, M.A.O., Scannell, J.W., Young, M.P. (2000) Anatomical connectivity defines the organization of clusters of cortical areas in the macaque monkey and the cat. Philos Trans R Soc Lond B Biol Sci 355, 91-110.
Holekamp, K.E. (2007) Questioning the social intelligence hypothesis Trends Cogn. Sci. 11, 65-69
Holzel, B.K., Ott, U., Hempel, H., Hackl, A., Wolf, K., Stark, R., Vaitl, D. (2007) Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neurosci Lett 421, 16-21.
Hoy, R.R. (1992) The evolution of hearing in insects as an adaptation to predation from bats, in Webster, D.B., Fay, R.R., Popper, A.N., eds.. The evolutionary biology of hearing. (Berlin Heidelberg New York: Springer), pp. 115129
Humphrey, N. (1976). The social function of intellect, in Growing Points in Ethology. Bateson, P.P.G., Hinde, R.A., eds. (Cambridge: Cambridge University Press) pp. 303-317.
Hurley S, Noe, A (2003) Neural plasticity and consciousness. Biology and Philosophy 18, 131-168.
Iwasaki, M., Delago, A., Nishino, H., Aonuma, H. (2006) Effects of previous experience on the agonistic behaviour of male crickets, Gryllus bimaculatus. Zoolog Sci 23, 863-872.
Jablonka E, Lamb, MJ (2005) Evolution in Four Dimensions (Cambridge, MA: MIT Press)
Jeong, H., Tombor, B., Albert, R., Oltvai, Z.N., Barabasi, A.L. (2000) The large-scale organization of metabolic networks. Nature 407: 651-654.
Kaku, M. (1996) Hyperspace (New York: Anchor)
Kauffman, S. (1993) The Origins of Order (New York: Oxford University Press)
Kolbert, E. (2006) Field Notes from a Catastrophe: Man, Nature and Climate Change (New York: Bloomsbury)
Lakoff, G. and Johnson, M. (1999) Philosophy in the Flesh: The Embodied Mind and its Challenge to Western Thought (New York: Basic)
Lazar, S.W., Kerr, C.E., Wasserman, R.H., Gray, J.R., Greve, D.N., Treadway, M.T., McGarvey, M., Quinn, B.T., Dusek, J.A., Benson, H., Rauch, S.L., Moore, C.I., Fischl, B. (2005) Meditation experience is associated with increased cortical thickness. Neuroreport 16, 1893-1897.
Lloyd, S. (2006) Programming the Universe (NY: Knopf)
Lutz, A., Dunne, J.D., Davidson, R.J. (2008). Meditation and the neuroscience of consciousness, in Cambridge Handbook of Consciousness. Zelazo, P., Moscovitch, M. Thompson, E., eds., (Cambridge: Cambridge University Press) pp. 497-549
Mayr, E. (1982) The Growth of Biological Thought: Diversity, Evolution and Inheritance (Cambridge, MA: Belknap)
McShea DW (1996) Metazoan complexity and evolution: is there a trend? Evolution 50: 477-492.
McShea DW (2005) The evolution of complexity without natural selection, a possible large-scale trend of the fourth kind. Paleobiology 31: 146-156.
McShea, D.W., Changizi, M.A. (2003) Three puzzles in hierarchical evolution. Integr Comp Biol 43: 74-81.
Metzger, M., Bernstein, C., Hoffmeister, T.S., Desouhant, E. (2010)Does Kin Recognition and Sib-Mating Avoidance Limit the Risk of Genetic Incompatibility in a Parasitic Wasp? PLoS One. 2010; 5(10): e13505
Milgram S (1967) The small world problem. Psychology Today 2, 60-67.
Mitchell, T.M., Hutchinson, R., Just, M.A., Niculescu, R.S., Pereira, F., Wang, X .(2003) Classifying instantaneous cognitive states from fMRI data. AMIA Annu Symp Proc, 465-469.
Monod, J. (1972) Chance and Necessity. An Essay on the Natural Philosophy of Modern Biology (New York: Vintage)
Newberg, A. (2002) Why God Won't Go Away. Brain Science and the Biology of Belief (New York: Ballantine)
Noe, A., Thompson, E. (2004) Are there neural correlates of consciousness? J Consciousness Studies 11, 3-28.
Orme-Johnson, D.W., Haynes, C.T. (1981) EEG phase coherence, pure consciousness, creativity and TM-Sidhi experiences. Int J Neurosci 13, 211-217.
Piaget, J. (1973) The Language and Thought of the Child (London: Routledge and Kegan Paul)
Pinker, S. (2007) The Language Instinct (New York: Harper Perennial)
Powell, R, ed. (2006) Talks With Ramana Maharshi: On Realizing Abiding Peace and Happiness (Carlsbad, CA: Inner Directions Foundation)
Prete, F.R .(1990) Configural prey recognition by the praying mantis, Sphodromantis lineola (Burr.); effects of size and direction of movement. Brain Behav. Evol 36, 300-306.
Prete, F.R. (1992) Discrimination of visual stimuli representing prey versus non-prey by the praying mantis Sphodromantis lineola (Burr.). Brain Behav. Evol 39. 285-288.
Preuss TM (2001). The discovery of cerebral diversity: an unwelcome scientific revolution, in Evolutionary Anatomy of the Primate Cerebral Cortex Falk, D., Gibson, K.R. eds. (Cambridge: Cambridge University Press) pp. 138-164.
Prothero, D. (2007) Evolution: What the Fossils Say and Why it Matters (NY: Columbia Univ. Press)
Raff, R.A. (1996) The Shape of Life (Chicago: University of Chicago Press)
Ramachandran, V.S., Hirstein, W. (1997) Three laws of qualia: what neurology tells us about the biological functions of consciousness. J. Consciousness Studies 4, 429-457
Rankin, C.H. (2005) Nematode memory: now, where was I? Curr. Biol. 15, R374-375.
Reader, S.M., Laland, K.N. (2002) Social intelligence, innovation, and enhanced brain size in primates Proc. Natl. Acad. Sci USA 99,4436-4441
Rizzolatti, G., Craighero, L. (2004) The mirror-neuron system. Annu. Rev,. Neurosci. 27, 169-192.
Schneider, R.A.Z., Schneider, R.W.S., Moore, P.A. (1999) Recognition of dominance status by chemoreception in the red swamp crayfish, Procambarus clarkii J Chem Ecol 25, 781-794.
Seager, W. (1999) Theories of Consciousness (New York: Routledge)
Searle, J.R. (1992) The Re-Discovery of the Mind (Cambridge, MA: MIT Press)
Serio, T.R., Lindquist, S.L. (2000) Protein-only inheritance in yeast: something to get [PSI+]-ched about. Trends Cell Biol 10, 98-105.
Shinkareva, S.V., Mason, R.A., Malave, V.L., Wang, W., Mitchell, T.M., Just, M.A. (2008) Using fMRI brain activation to identify cognitive stages associated with perception of tools and dwellings. PLoS ONE(1), 1-9.
Simon, H. (1962) The architecture of complexity. Proc Am Phil Soc 106, 467-482.
Smith, A.P. (2009) The Dimensions of Experience (Xlibris)
Smolin, L. (2006) The Trouble With Physics: The Rise of String Theory, The Fall of a Science, and What Comes Next (New York: First Mariner)
Sober, E., Wilson, D.S. (1998) Unto Others. The Evolution and Psychology of Unselfishness (Boston, MA: Harvard University Press)
Sporns, O., Zwi, J.D. (2004) The small world of the cerebral cortex. Neuroinformatics 2, 145-162.
Thompson, E., Varela, F.J. (2001) Radical embodiment: neural dynamics and consciousness. Trends Cogn Sci 5: 418-425.
Tibbetts, E.A. (2002) Visual signals of individual identity in the wasp Polistes fuscatus. Proc R Soc Lond B Biol Sci 269, 1423-1428.
Tononi, G., Sporns, O,, Edelman, G. (1994) A measure for brain complexity: relating functional segregation and integration in the nervous system. Proc. Natl. Acad. Sci USA 91, 5033-5037.
Valentine, J.W. (2003) Architectures of biological complexity. Integr Comp Biol 43, 99-103.
Van der Velden, J., Zheng, Y., Patullo, B.W. Macmillan, D.L. (2008) Crayfish Recognize the Faces of Fight Opponents PLoS ONE 3, e1695
Walls, M.L,, Layne, J.E. (2009) Fiddler crabs accurately measure two-dimensional distance over three-dimensional terrain. J Exp Biol. 212, 3236-40.
Watts, D.J, Strogatz, S.H. (1998) Collective dynamics of small world networks. Nature 393, 440-442
Whiten, A. (2007) Pan-African culture: memes and genes in wild chimpanzees. Proc. Natl. Acad Sci. USA 104, 17559-17560
Wickner, R.B., Taylor, K.L., Edskes, H.K., Maddelein, M.L., Moriyama, H., Roberts, B.T. (2000) Prions of yeast as heritable amyloidoses. J Struct Biol 130, 310-322.
Wilber, K. (1995) Sex, Ecology, Spirituality (Boston, MA: Shambhala)
Wilkins (1997) Evolution and Philosophy. A Good Tautology is Hard to Find http://www.talkorigins.org/faqs/evolphil/tautology.html
Wilson, E.O. (2007) The Creation: An Appeal to Save Life on Earth (New York: Norton)
Wohlgemuth, S., Ronacher, B., Wehner, R. (2001) Ant odometry in the third dimension.
Nature 411, 795-8
Wright, R. (2001) Nonzero: The Logic of Human Destiny (New York: Vintage)
Wuchty, S. (2001) Scale-free behavior in protein domain networks. Mol. Biol. Evol. 18: 1694-1702.
Yook, S.H., Jeong, H., Barabasi, A.L. (2002) Modeling the Internet's large-scale topology. Proc Nat Acad Sci USA 99, 13382-113386.
|
 Andrew P. Smith, who has a background in molecular biology, neuroscience and pharmacology, is author of e-books Worlds within Worlds and the novel Noosphere II, which are both available online. He has recently self-published "The Dimensions of Experience: A Natural History of Consciousness" (Xlibris, 2008).
Andrew P. Smith, who has a background in molecular biology, neuroscience and pharmacology, is author of e-books Worlds within Worlds and the novel Noosphere II, which are both available online. He has recently self-published "The Dimensions of Experience: A Natural History of Consciousness" (Xlibris, 2008).