|
TRANSLATE THIS ARTICLE
Integral World: Exploring Theories of Everything
An independent forum for a critical discussion of the integral philosophy of Ken Wilber
 Federico Nicola Pecchini is an independent researcher and activist focusing on conscious evolution, integral development, cooperative economics and transnational sociology. He is currently based in Dharamsala, India. Contact: fpecchini @ gmail.com. Federico Nicola Pecchini is an independent researcher and activist focusing on conscious evolution, integral development, cooperative economics and transnational sociology. He is currently based in Dharamsala, India. Contact: fpecchini @ gmail.com.
 What is Consciousness?Natural Religion, Part 4Federico Nicola Pecchini
“Biologically, we are just another ape. Mentally, we are a new phylum of organisms.”
—Terrence Deacon As more regularization processes got synergistically coupled together, selves and their ability to regenerate evolved to more efficient forms. And as they started to interact with with each another and got synergistically coupled between themselves, tangled hierarchies of teleodynamic processes started to emerge, which make up complex organisms as we see today. Deacon and his team acknowledge that the minimal autogen model described before satisfies only the very basic conditions for a teleodynamic process to occur (self-regeneration, protection and reproduction) and thus should be regarded as a workable hypothesis addressing the missing link between life and non-life (abiogenesis). To bridge the gap with present life, what is left to explain is the emergence of its now ubiquitous semi-permeable membrane (which achieves selective interaction with the environment) and of its highly refined self-replicating genome (which achieves internal representation in the form of a functional set of instructions that can be copied and passed on).  Terrence Deacon At first, the authors imagine that the minimal autogen would be continuously contaminated by other molecules which could impair autocatalysis. In most cases, the autogen would be “threatened by an accumulation of deleterious molecules that […] would reduce its capacity for self-regeneration”, but in some cases it could also end up incorporating some beneficial catalyst which makes the whole cycle more efficient. Those autogen lineages would have increased capacity for self-regeneration and reproduction, and would thus be favored by natural selection. Then they imagine an “autogen lineage that through catalyst substitution produced a variation of the original capsid that opens more readily in the presence of reactants”. Such an evolved autogen may be called “selective”, since it would “have internalized a capacity for true selective interaction”, and thus even greater self-regenerative efficiency. Finally, they imagine an autogen which “also produces energy-ferrying nucleotide by-products that capture energy in the form of phosphates and transfer it to other catalytic processes”. In the closed phase, the nucleotides would polymerize to form a double helix and the autogen’s catalysts would tend to adhere to its surface. But once the autogen breaks open, the catalyst-release sequence would be determined by their disposition on the helix and thereby the most efficient release sequences (or “templates”) would again be naturally selected.  Jeremy Sherman Sherman writes that “the autogen is an interpretation, a bet about what might work given or about its environment”. The minimal autogen interprets its environment directly, by means of its emergent dynamics. At this stage “we can’t distinguish between sign and self”, hence Deacon calls it “proto-iconic”. “With the selective autogen, we can distinguish a one sign relationship. A reactant attaching to the closed capsid surface is interpreted as a sign to open [and thus] the surface becomes something like a simple sense organ”. Deacon calls this interpretive competency “proto-indexical” or proto-sentient. “With template autogens, signs become internalized as a distinct static part of the self. […] The template sequence is a record of past success in the form of functionally constrained sequences.” He calls this third kind of interpretation “proto-symbolical”, “because a system of nucleotide sequences has beecome adaptively correlated to a system of catalyst release sequences.” Sherman continues: “With the template autogen, there is a loose evolutionary parallel to the aha moment we each have in childhood, or more importantly the aha moment that happened when humankind discovered the power of language. […] The template sequences provide a static representation of a key aspect of the autogen’s dynamic. They become a mechanism for constraining error catastrophe, a means for orchestrating catalyst release. Having this heritable advantage provides autogens with a static material medium upon which other functional constraints might accumulate. […] The template thus provides a redundant medium for protecting and reproducing functional constraints. The template is redundant in that it re-presents the autogen’s dynamics. Thus the template autogen carries forward two copies of the same constraints, one in the autogen’s dynamic alteration between open and closed parts, the other in the static template molecule sequences.” In Shannon’s information theory, “information” is measured as the divergence from expectations about what’s likely to occur in the future. Redundancy, as in repeating a message over and over again, increases communication reliability over background noise. “In the template autogen, the protection function of self-regeneration becomes partially offloaded onto the template, allowing for greater evolved variability”, since “redundancy relaxes the constraints on the dynamics, tolerating greater variance and making it possible to accumulate a wider range of adaptations”. If the new mutations or combinations of nucleotide sequences prove to be not functional to the survival of the self, they’ll be lost thanks to natural selection. Otherwise, the new instructions will be successfully carried forward from generation to generation, inscribed in the code of a self’s lineage (speciation). The template sequence is thus a record of past success in the game of life, and represents for the first living beings an evolvable set of inherited expectations about the right way to interact with their environment. It is an inbuilt capital of practical knowledge (know-how), which provided our ancestors with a secure base to experiment with new functional adaptations.

Three Sphinxes of Bikini—Salvador Dali
That’s how life was able to “laboriously carve out degrees of causal freedom from the fabric of existence” (see free will). And as template sequences became more redundant, this margin of freedom-within-constraints also increased. Living beings had more options to “choose” from. Obviously at first it was not a subjective choice but rather a function of the self’s accumulated constraints. But nonetheless we can argue that even the genetic template of simple prokaryotic cells—just like the nervous system at later stages of biological evolution—already functions as an ‘anticipation machine’ that guides the self in its explorative interpretation of the world, and should then come to be regardeded as an emergent proto-mind or proto-consciousness (with consciousness broadly defined here as “self-represented awareness”). Here’s a proposed nested hierachy of teleodynamic systems:
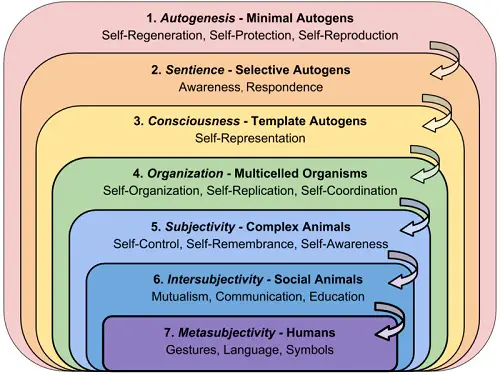
The emergence and evolution of living consciousness
In the last phase of the Proterozoic—about a billion years ago—these single- celled bacteria then merged into multicellular organisms, where each cell learned to perform a specialized function within a larger body. As eukaryotes, fungi and plants covered the surface of the planet, they set the conditions for more complex life to emerge: half a billion years later—with the Cambrian explosion—most of the known animal species suddenly appeared on record. As their nervous system developed, these new complex organisms learned to condition their instinctive behavior according to their own subjective experiences (eg. memory of success or failure), thus overriding the genetic template with a psychic template. Those who lived in groups then learned to pass on their subjective knowledge to their offspring and fellow specimen. We can observe how, in certain classes of highly evolved social animals such as birds or mammals, individuals are capable of communicating to others what they learn by direct experience during their life. Such knowledge is no longer passed on via the genes, but in the guise of a cultural tradition. Hence the new generations can learn, by imitating the behavior of the adults, how to recognize a certain specific threat or how to perform a complex skill set. Eventually, only a few hundred thousand years ago (a blink of an eye in the timespan of biological evolution), we get to us humans and to our unique combination of symbolic thought and discursive language—which allowed us to talk about things also if not immediately present as we spoke, by mentally representing them as concepts and ideas within our imagination. Cognitive neuroscientists call this feature Mental Time Travel (MTT), since it involves the capacity to reconstruct events from the past (episodic memory) as well as to imagine possible scenarios in the future. This new outstanding ability—further enhanced by the invention of writing—enabled the creation of that immensely articulated and multi-generational cultural heritage which is the also the most striking and unique characteristic of our species. In his first book, Deacon had appropriately called us “The Symbolic Species”: “Though we share the same earth with millions of living creatures, we also live in a world that no other species has access to.
Human consciousness came forth as the psychic wave emerging from the genetic memory folded on itself and, recognizing its own image in the mirror of self-awareness, started questioning itself about the meaning of existence.
Our storytelling species was the first, in life’s long evolutionary journey, to become aware of the same evolutionary processes from which it was born. In a sense, we could really say that “we are the universe becoming conscious of itself”. But at this point we must be more specific: human consciousness is not an inherent property of the entire universe, nor is it a gift from some supernatural God. Rather, it emerged right here on Earth, rooted in the blood of our brothers animals and the lymph of our sisters plants, in this familiar and homely environment we call Nature or biosphere. Our species has grown beyond the planetary boundaries and today is even sending probes outside the solar system, but the only thing we get by aiming too far into space is a distinctive feeling of existential vertigo. That peak of human consciousness alluded to by so many spiritual traditions in the past is not some kind of loose cosmic feeling where we can ideally attain eternal rest and lose all attachments to the world, but is really that moment when the biosphere completes a U-turn on its axis and is able to see itself from outer space, becoming at once aware of its common past and responsible for its common future. Our roots are here—on Earth—with the multifaceted community of life. If we cut our roots we are just an insignificant speck lost in the void. If instead we turn back towards Nature, embrace our roots and nurture them, we’ll feel empowered and infused with living energy, ready to face whatever challenge the universe may be throwing at us. To make good use of our highly evolved consciousness we must accept and recognize our natural origins and limitations. By becoming aware both of our existential precariousness and our synergistic potential, we can learn to face the world more wisely and responsibly, as a fully mature metasubjective species. We can learn to transcend and include the individual self in that larger Self which is the evolutionary journey of life itself. As Krishna said: “He who experiences the unity of Life sees his own self in every living being, and every living being in his own self." FURTHER READINGTerence Deacon, The Symbolic Species: The Co-evolution of Language and the Brain, W. W. Norton & Company, 1998. Terence Deacon, Incomplete Nature: How Mind Emerged from Matter, W. W. Norton & Company, 2011. Jeremy Sherman, Neither Ghost nor Machine: The Emergence and Nature of Selves , Columbia University Press, 2017.
|